Растителни тъкани
Клетките са асоциирани заедно по различен начин и формират структурни и функционални единици, наричани тъкани.
Растителните тъкани представляват съвкупност от свързани помежду си клетки, които:
- имат общ произход,
- еднаква структура и
- изпълняват една и съща функция.
Според основното си предназначение, тъканите се подразделят на:
- Меристемни (образувателни) тъкани - обслужват нарастването на растенията
- Трайни тъкани - изпълняват всички останали функции
Трайните тъкани са организирани в по-големи единици, известни като тъканни системи
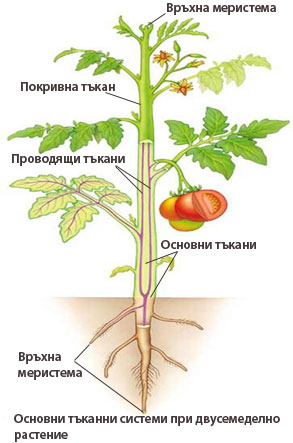
Тъканните системи се срещат във всички органи на растението, като преминават без прекъсване от орган в орган, обуславяйки единността и сходството в строежа на растителното тяло.
Растенията имат следните тъканни системи:
- Покривна тъканна система - изгражда най-външния защитен слой на растителното тяло и е представена от епидермис и корк (при наличие на вторично нарастване на растителния орган).
- Проводяща тъканна система - състои се от два вида проводящи тъкани (ксилема и флоема) и обслужва транспорта на веществата в растението. Потопена е сред основните тъкани
- Основна (фундаментална) тъканна система - обслужва всички останали функции на растението и се състои от паренхима, склеренхима и коленхима. Паренхимата е най-широко застъпената основна тъкан.
Диференцирането на горепосочените тъканни системи се инициира още при развитието на зародиша. Техни предшественици са първичните меристеми на зародиша - протодерма, прокамбий и основна меристема, съответно.
В растителното тяло, отделните тъканни системи са разположени по типичен за дадения орган и таксон начин
Обикновено разположението на тъканните системи в отделните растителни органи е сходно. Като правило проводящите тъкани са потопени сред основните, а покривните тъкани формират най-външния слой на растителните органи. Принципните различия идват най-вече от относителното разположение на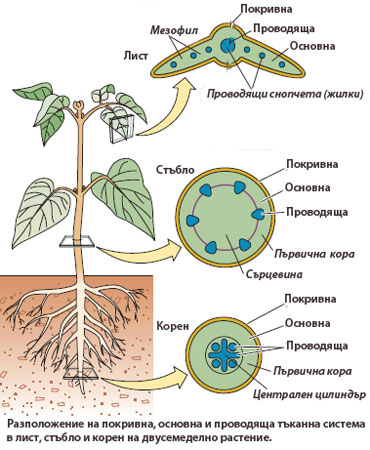 проводящите и основните тъкани една спрямо друга.
проводящите и основните тъкани една спрямо друга.
Например, в стъблото на двусемеделните растения, проводящите снопчета са подредени в кръг и са потопени сред основните тъкани. Зоната оставаща навътре от проводящите снопчета се нарича сърцевина, а зоната навън от тях - първична кора.
В корена на същото растение проводящите тъкани формират централен цилиндър, обграден от първична кора.
В листа проводящата тъканна система е представена от проводящи снопчета (жилките) потопени сред фотосинтезираща паренхима (мезофил).
В зависимост от това, дали са изградени от един, два или повече типа клетки, тъканите се подразделят на:
- прости тъкани - изградени са от един тип клетки (паренхима, коленхима, склеренхима)
- сложни (комплексни) тъкани - изградени са от два или повече типа клетки (ксилема, флоема, епидерма)
Според произхода си тъканите се подразделят на:
- първични - водят началото си от първични меристеми
- вторични - водят началото си от вторични меристеми
Повечето от тъканите са многофункционални и изпълняват две или няколко функции. Освен това, тъканите могат да променят функциите си или да запазят само някоя от първоначалните. Например дървесината първоначално има преди всичко проводяща функция, която впоследствие губи и служи само за механична опора на растението.
Меристемни (образувателни) тъкани
След поникването, формирането на тялото на растението зависи изцяло от дейността на меристемните тъкани (меристемите). Терминът меристема (от гръцки merismos - "делене") дефинира деленето на клетките, като основна функция на меристемните тъкани.
Меристемните тъкани обуславят нарастването на растителното тяло. Клетките им се делят, образуват нови клетки, които увеличават размерите си и се диференцират, формирайки всички растителни тъкани и органи.
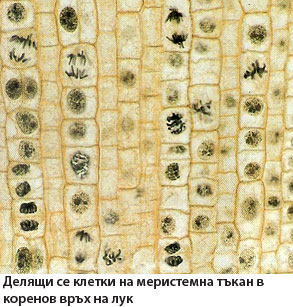 Меристемните тъкани на растенията запазват способността си да се делят неопределено дълго време, с което обуславят нарастването и образуването на нови органи през целия живот на растенията.
Меристемните тъкани на растенията запазват способността си да се делят неопределено дълго време, с което обуславят нарастването и образуването на нови органи през целия живот на растенията.
Клетките на меристемните тъкани са:
- живи,
- дребни,
- изодиаметрични,
- без междуклетъчни пространства,
- с тънки клетъчни стени,
- относително голямо ядро и
- гъста цитоплазма
→ богата на свободни рибозоми;
→ има слабо развити ендоплазмена мрежа и апарат на Голджи;
→ пластидите са представени от пропластиди
Меристемните тъкани се състоят от инициални клетки и техни производни
- Инициални клетки - делят се неограничен брой пъти, като винаги едната от новополучените клетки при деленето остава инициална, а втората е производна меристемна клетка;
- Производни меристемни клетки - делят се определен брой пъти, след което започват да увеличават размерите си и да се диференцират в клетки на трайните тъкани.
Растенията могат да имат във връхната си меристема:
- една инициална клетка - мъхове и папрати; или
- група от инициални клетки - голосеменни и покритосеменни растения
Според произхода си меристемите се подразделят на първични и вторични
- Първични меристеми - водят началото си от зародиша на семето (връхни меристеми, перицикъл).
→ Тъканите образувани от дейността на първичните меристеми се наричат първични растителни тъкани.
→ Органите на растението изградени от първични тъкани имат първичен анатомичен строеж.
→ Нарастване включващо формиране на първичните растителни тъкани се нарича първичен растеж.
- Вторични меристеми - възникват по-късно в живота на растението и водят началото си от някои от трайните тъкани (най-често от паренхимните), които се дедиференцират и отново придобиват способност да се делят (камбий, фелоген).
→ Тъканите образувани от вторичните меристеми се наричат вторични тъкани.
→ Органите на растението изградени от вторични тъкани имат вторичен анатомичен строеж.
→ Нарастване включващо формиране на вторични растителни тъкани се нарича вторичен растеж.
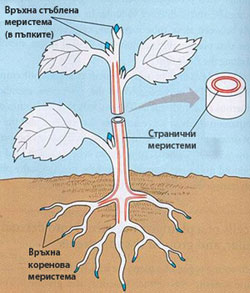 Според месторазположението си меристемите се подразделят на:
Според месторазположението си меристемите се подразделят на:
- Връхни меристеми - разположени са по върховете на корена и стъблото и разклоненията им;
→ обуславят нарастването на дължина на корените и стъблата; - Странични меристеми - разположени са странично в осевите органи на растението;
→ осигуряват надебеляването на корена и стъблото при храсти, дървета и някои тревисти растения; - Интеркаларни (вмъкнати) меристеми - вмъкнати са сред повече или по-малко диференцирани тъкани; остатък са от връхните меристеми и действат на известно разстояние от тях;
→ осигуряват удължаване на дадения растителен орган (например стъблата на житните треви)
Връхни (апикални) меристеми и техните производни
Разположени са по върховете на корена и стъблото и разклоненията им и обуславят нарастването на растителните органи на дължина. Водят началото си от меристемните клетки на зародиша и са първични по произход.
Клетките на връхните меристеми не са равностойни
Във връхните меристеми инициалната клетка или групата инициални клетки заедно с най-близко лежащите до тях производни клетки се обозначават като протомеристема (промеристема). Клетките на протомеристемата имат способността да се делят неограничен брой пъти и запазват меристемния си характер да края на живота на растението. Те съответстват на стволовите клетки при човека.
Деленето на клетките, обаче не е ограничено само в зоната на връхната меристема. То протича и в по-отдалечените и частично диференцирани меристемни тъкани, известни като първични (транзиционни) меристеми. Тези транзиционни меристеми - протодерма, основна меристема и прокамбий - се образуват от дейността на връхната меристема и формират цилиндрични слоеве.
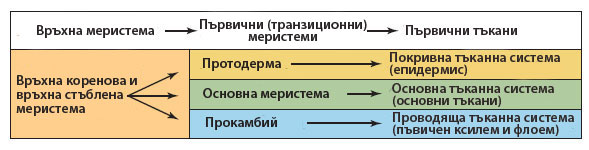
Клетките на първичните (транзиционните) меристеми остават меристемни за известно време, след което, основно според месторазположението си, се диференцират в клетки на трите тъканни системи - покривна, основна и проводяща, като:
- от протодермата се образува епидермата,
- от прокамбия - първичните проводящи тъкани, а
- от основната меристема - всички останали основни тъкани (паренхима, коленхима, склеренхима)
Връхни коренови меристеми
Образуват нови клетки и допринасят за нарастването на корените и разклоненията им на дължина, осигурявайки на растенията достъп до вода и хранителни вещества.
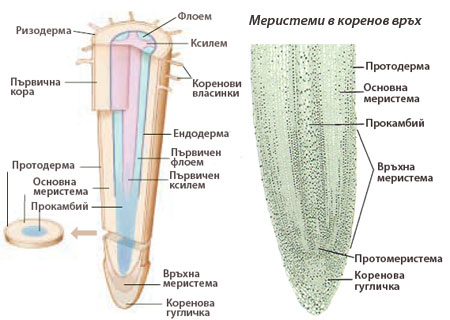 Част от производните на връхната коренова меристема образуват кореновата гугличка, която предпазва кореновите връхчета.
Част от производните на връхната коренова меристема образуват кореновата гугличка, която предпазва кореновите връхчета.- Останалите производните клетки на връхната коренова меристема много скоро се диференцират в клетки на първичните транзиционни меристеми протодерма, основна меристема и прокамбий, които формират ясно разграничими цилиндрични слоеве.
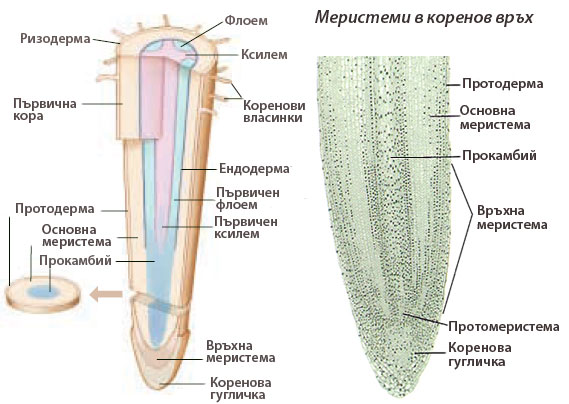
Впоследствие първичните транзиционни меристеми се диференцират в следните трайни тъкани:
- протодермата образува покривната тъкан на корена (ризодерма),
- основната меристема - тъканите на първичната кора, а
- прокамбия - проводящите тъкани на централния цилиндър.
Първоначално клетките на протомеристемата са много активни, но след време интензивността на деленето на клетките в тази зона намалява. Най-интензивно започват да се делят клетките на връхната меристема разположени непосредствено под клетките на протомеристемата.
Връхни стъблени меристеми
Разположени са във връхните и страничните пъпки на стъблото и клонките. Освен, че образуват нови клетки и осигуряват нарастването на дължина на стъблото и клонките, те още образуват и
- листни зачатъци (листни примордии) - които се развиват в листа, и
- зачатъци на странични пъпки - които се развиват в странични клонки
Връхната стъблена меристема е само тази част от вегетационния връх, разположена над най-младия листен зачатък, докато вегетационния връх включва връхната стъблена меристема и субапикалната зона носеща млади листни зачатъци.
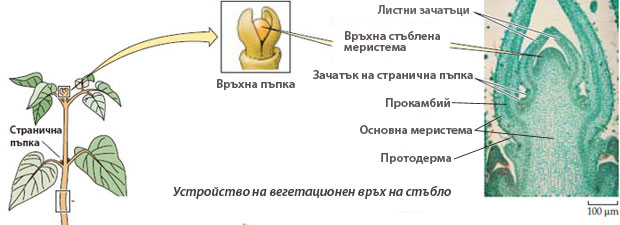
Връхната стъблена меристема при повечето цветни растения има т.н. туника-корпус организация
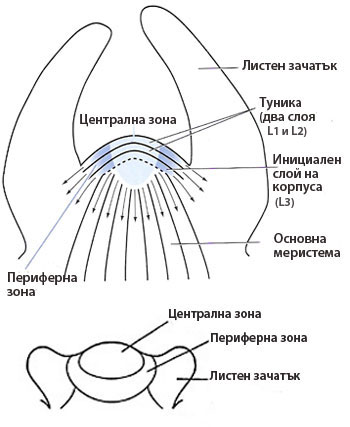 Във връхната стъблена меристема се разграничават две зони - туника и корпус, които се различават по плоскостта в която се делят клетките им.
Във връхната стъблена меристема се разграничават две зони - туника и корпус, които се различават по плоскостта в която се делят клетките им.
- Туника
→ състои се от най-външния слой или слоеве клетки на връхната меристема;
→ клетките на туниката се делят антиклинално (перпендикулярно на повърхността на органа);
→ осигурява повърхностно нарастване, без да се увеличават меристемните слоеве;
→ дава началото на епидермата;
→ цветните растения най-често имат туника от два слоя клетки, обозначавани като L1 и L2 (L1 е външния слой). - Корпус
→ състои се от останалите клетки лежащи под туниката;
→ инициалните клетки на корпуса са организирани в един слой, разположен непосредствено под вътрешния слой на туниката (обозначава се като L3);
→ инициалните клетки на корпуса се делят периклинално (успоредно на повърхността на органа);
→ останалите клетки на корпуса се делят във всички посоки;
→ от клетките на корпуса се диференцират тъканите на първичната кора и централния цилиндър на стъблото;
Корпусът и всеки слой на туниката имат собствени инициални клетки, като инициалните клетки на корпуса се разполагат непосредствено под тези на туниката.
На върха на вегетационния конус на цветните растения корпусът и туниката участват в обособяването на две зони:
- централна зона - обособява се от централните зони на инициалните слоеве на туниката и корпуса и съответства на протомеристемата на връхната стъблена меристема, и
- периферна зона - обособява се от периферните зони на инициалните слоеве на туниката и корпуса и обгражда централната зона, образувайки пръстен около нея.
Периферната зона е митотично много активна, докато деленето на клетките в централната зона не е особено интензивно.
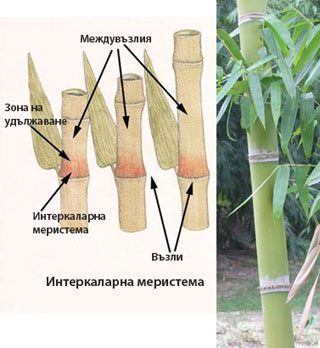 Интеркаларни (вмъкнати) меристеми
Интеркаларни (вмъкнати) меристеми
Интеркаларните меристеми:
- са остатък от връхните меристеми
- действат на известно разстояние от тях
- вмъкнати са сред вече диференцирани тъкани
- действат за кратък период от време
- осигуряват удължаване на дадения растителен орган
Причината за кратковременната дейност на интеркаларните меристеми е липсата на инициални клетки при тях.
Интеркаларни меристеми има в основата на междувъзлията и листните влагалища на житните треви, в основата на плодниците (фъстък) и др. Освен за удължаване на междувъзлията, вмъкната меристема спомага и за изправяне на стъблата на житните при полягане.
Странични меристеми
Камбий и фелоген (корков камбий) са примери за странични меристеми. Те са вторични по произход и осигуряват вторично надебеляване на осевите органи на повечето храсти и дървета, а също и на някои тревисти растения. Представляват цилиндри от меристемни тъкани, разположени успоредно на дългата ос на стъблата и корените.
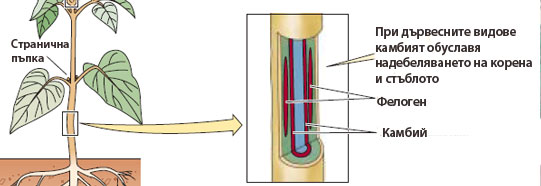
Камбият образува вторичните проводящи тъкани - вторична дървесина и вторично лико
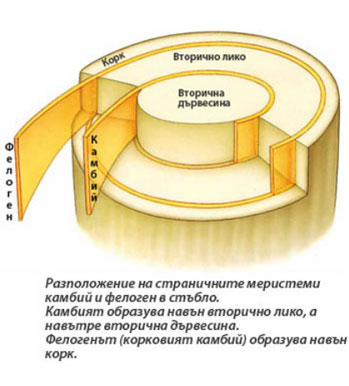 Камбият представлява цилиндър от няколко слоя клетки - един слой инициални клетки и техните най-близки производни.
Камбият представлява цилиндър от няколко слоя клетки - един слой инициални клетки и техните най-близки производни.- Състои се от два вида клетки -
→ вретеновидни инициали - удължени и заострени в краищата си клетки и
→ лъчеви инициали - почти изодиаметрични дребни клетки. - Клетките на камбия се делят тангенциално (= на периклинално → успоредно на повърхността на органа) и клетките отделени навън от камбиалния пръстен се диференцират в клетки на вторичното лико, а тези отделени навътре - в клетки на вторичната дървесина.
- Клетките на камбия са вторични по произход и произлизат от клетки на прокамбия в проводящите снопчета и дедиференцирани перенхимни клетки разположени между проводящите снопчета.
Фелогенът (корковият камбий) образува вторичната покривна тъкан - корк
- Залага се в първичната кора на корена и стъблото
- Като правило, първият заложен фелоген винаги представлява цилиндър от няколко слоя тънкостенни клетки с правоъгълна форма.
- Клетките му се делят тангенциално и навън образуват многобройни слоеве коркови клетки, а навътре - няколко слоя клетки на фелодермата.
 Вторичен по произход и произлиза от дедиференцирани клетки на епидермата или паренхимни клетки на първичната кора.
Вторичен по произход и произлиза от дедиференцирани клетки на епидермата или паренхимни клетки на първичната кора.
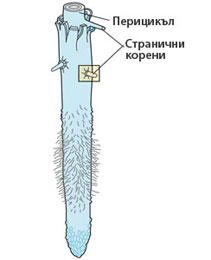
Перицикълът образува страничните разклонения на корена
- Перицикълът е първична странична меристема.
- Формира цилиндър от един или няколко слоя меристемни клетки.
- Формира първия слой на централния цилиндър на корена.
- Образува страничните разклонения на корена.
- Първичен е по произход - води началото си от прокамбия.
 Ранева меристема
Ранева меристема
- Вторична по произход - възниква при нараняване на органите на растенията.
- Клетките около мястото на нараняването се меристемизират, делят се и образуват коркова тъкан, която покрива раната.
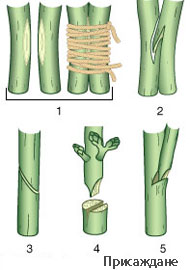
- Когато нараняването е дълбоко и прекъсне проводящите елементи, камбият и живите клетки около него участвaт в образуването на регенерационна тъкан, известна като калус.
- Калус се образува и при присаждането при лозата и другите овощни видове.
- На мястото на присаждането, от клетките на калуса се образуват всички тъкани, които свързват подложката с присадника (включително и проводящите тъкани).
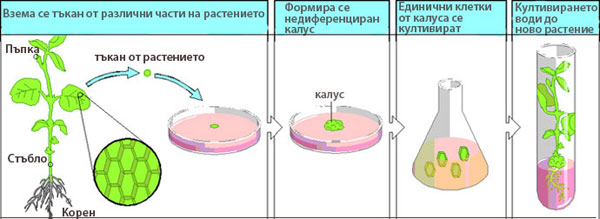
При тъканните култури също се образува калус, като от единични клетки на калуса, при подходящи условия, се регенерира цяло растение
Регенерационна меристема
При някои растения меристемни клетки могат да възникнат от клетките на епидермата или от власинките по нея, които регенерират цяло растение. Наблюдава се при бегония, фикус и др.

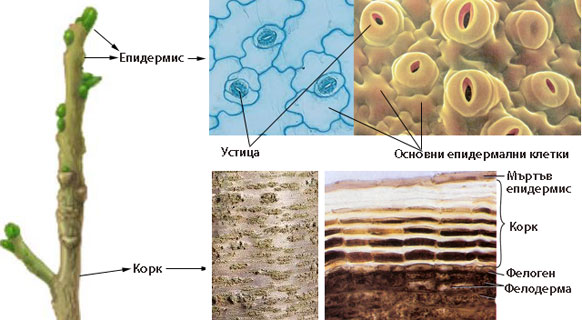
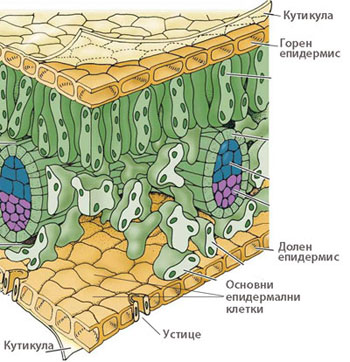
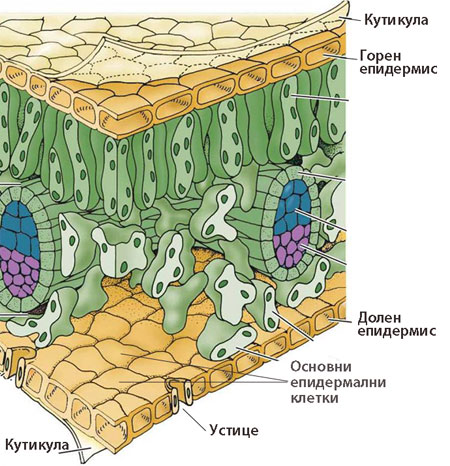
Изграждат най-външния, защитен слой на растителното тяло и предпазват растителните органи от неблагоприятните условия на външната среда. Представени са от:
- Епидермис - покрива надземните части на растението с първично анатомично устройство
- Ризодермис - покрива корените с първично анатомично устройство
- Корк (перидермис) - покрива растителните органи при наличие на вторично нарастване
Епидермисът покрива всички първично устроени растителни органи
 Епидермисът е покривната тъкан на листата, цветните части, плодовете, семената, а също и на стъблата, преди те да претърпят значително вторично нарастване.
Епидермисът е покривната тъкан на листата, цветните части, плодовете, семената, а също и на стъблата, преди те да претърпят значително вторично нарастване.- Първичен е по произход и води началото си от протодермата.
- При растенията без вторично нарастване епидермисът се запазва като покривна тъкан до края на живота им.
- При растенията с вторично нарастване, епидермисът се замества от вторична покривна тъкан - корк.
 Основни функции:
Основни функции:
- предпазва растението от неблагоприятните условия на околната среда
- регулира транспирацията и газообмена
- възпрепятства проникването на микроорганизми в тялото на растението
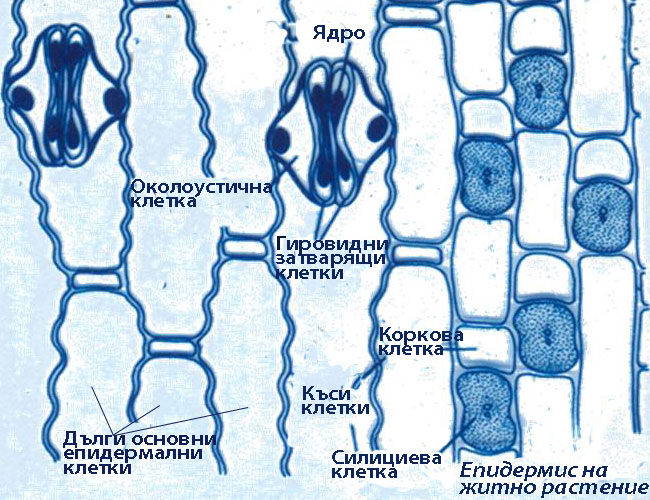
При повечето растения епидермисът е изграден е от един слой клетки, различаващи се по своята структура и функция.
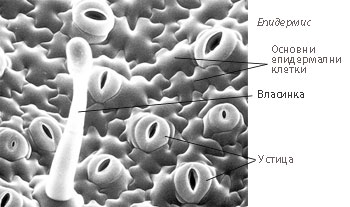
Епидермисът е комплексна тъкан и включва следните основни типове специализирани клетки:
 Основни епидермални клетки
Основни епидермални клетки- Устица
- Власинки
Основни епидермални клетки
- Те са най-многобройни и най-слабо диференцирани
- Живи клетки с разнообразна форма (обикновено плоски, с малка дълбочина и голяма външна и вътрешна повърхност), често с нагънати антиклинални стени.
- Разположени са компактно, без междуклетъчни пространства - придават значителна механична устойчивост.
- Имат голяма централна вакуола и обикновено са безцветни - хлоропластите са малко, с недоразвита тилакоидна система → съответно съдържат недостатъчно хлорофил
- Цитоплазмата е с добре развити ендоплазмена мрежа и апарат на Голджи
- Често външните стени на епидермалните клетки са надебелени и покрити с кутикула, която е почти напълно непропусклива за вода и газове.
Кутикула
- Представлява слой от кутин в който са вградени восъци (интракутикулярни восъци) и пектинови вещества (обуславят частичната й водопропускливост)
- Често е покрита и от повърхностен слой от восъци (епикутикулярни восъци). Те се отлагат под формата на зрънца, люспи, кукички, пръчици и др. и формират светлосив или синкав восъчен налеп (плодовете на слива, грозде, листа на зеле, перуника, лале и др.).
- Дебелината й варира, като растенията от сухите месообитания имат по-силно развита кутикула.
- При някои ксерофитни и сукулентни растения, восъците обграждат устицата и формират над тях сложни структури известни като "восъчни комини"
Функция:
- Предотвратява загубата на вода и други молекули от повърхността на растителните органи
- Предпазва растенията от развитието на бактериални и гъбни заболявания → не позволява задържането на вода и съответно прорастването и развитието на патогени върху растителните органи.

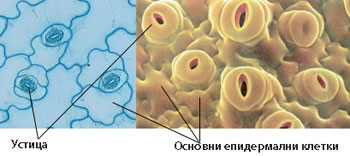
 Устица
Устица
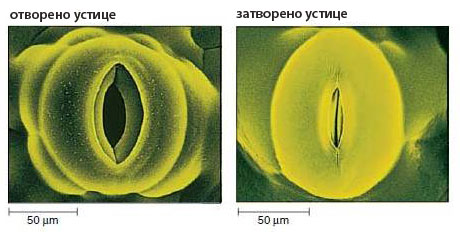
- Представляват отвори в епидермиса и регулират транспирацията и газообмена.
- Най-многобройни са в листата - от 1000 до 1 000 000 устица на cm2
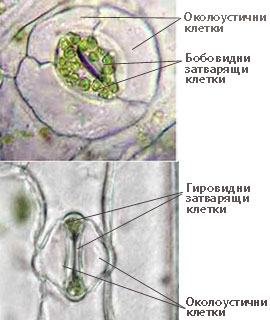
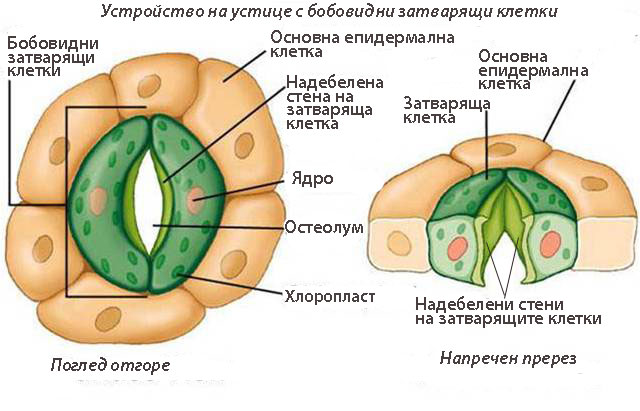
- Образувани са от две високо специализирани затварящи клетки, допрени с краищата си така, че помежду им се образува отворче - остеолум.
- При много растения около устицата има една или повече клетки, които се различават от основните епидермални клетки по форма, размери и разположение и се наричат околоустични клетки.
- Околоустичните клетки заедно със затварящите клетки на устицата образуват устичен (стоматален) апарат
Устройство на устицата
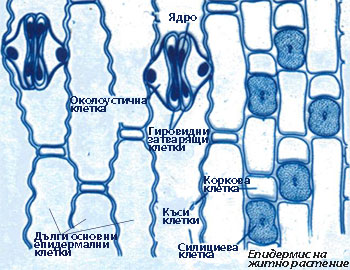 Затварящите клетки са с бобовидна или гировидна (при житните растения) форма.
Затварящите клетки са с бобовидна или гировидна (при житните растения) форма.- Клетъчната обвивка на затварящите клетки е неравномерно надебелена - надебелена към остеолума и тънка и еластична в останалите зони.
- Затварящите клетки имат голяма централна вакуола, добре развита ендоплазмена мрежа, апарат на Голджи, множество митохондрии, скорбяла съдържащи хлоропласти.
- Отварянето и затварянето на устицата регулира транспирацията и газообмена и се повлиява от фактори на околната среда като светлина, температура, достатъчна осигуреност с вода и СО2.
- Отварянето и затварянето на устицата е свързано с промяна на тургорното налягане в затварящите клетки - повишаването на тургорното налягане води до издуване на тънките стени на двете затварящи клетки, които изтеглят след себе си надебелените участъци и отварят устицето.
 Власинки
Власинки
При голям брой растения епидермисът образува разнообразни типове власинки, имащи най-вече защитна функция. Власинки се образуват по всички органи на растението и се характеризират с голямо морфологично разнообразие. Обикновено се развиват от деленето на една епидермална клетка.
Власинките са образувания на епидермата и се подразделят на покривни и жлезисти.
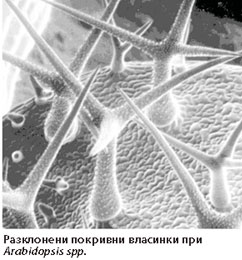
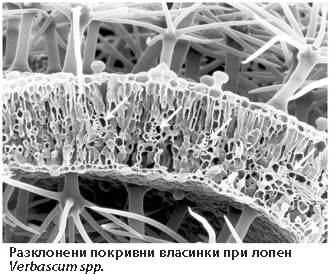 Покривни власинки
Покривни власинки
Имат защитна функция като:
- Отразяват слънчевата светлина, което понижава температурата на повърхността на листа
- Намаляват загубата на вода, като намаляват скоростта на движение на въздуха около устицата
- Осигуряват защита срещу насекоми - при много видове повишената овласеност корелира с повишена устойчивост на нападения от насекоми
- Осигуряват защита срещу тревопасни животни, като влошават вкусовите качества на растителните органи.
- Много епифити използват власинките по листата си за поглъщане на вода.
Покривните власинки се подразделят на:
 Неразклонени власинки - линейни власинки, могат да са едноклетъчни или многоклетъчни; многоклетъчните биват едноредови или многоредови.
Неразклонени власинки - линейни власинки, могат да са едноклетъчни или многоклетъчни; многоклетъчните биват едноредови или многоредови.- Разклонени власинки - те са многоклетъчни, храстовидни, дисковидни, звездовидни и др.
- Прикрепващи - линейни власинки, кукесто, Г-образно или Т-образно извити на върха.
- Коренови власинки - едноклетъчни, участват във всмукването на вода от почвата
- Папили - малки подутини на външните стени на епидермалните клетки, → някои автори не ги отнасят към власинките.
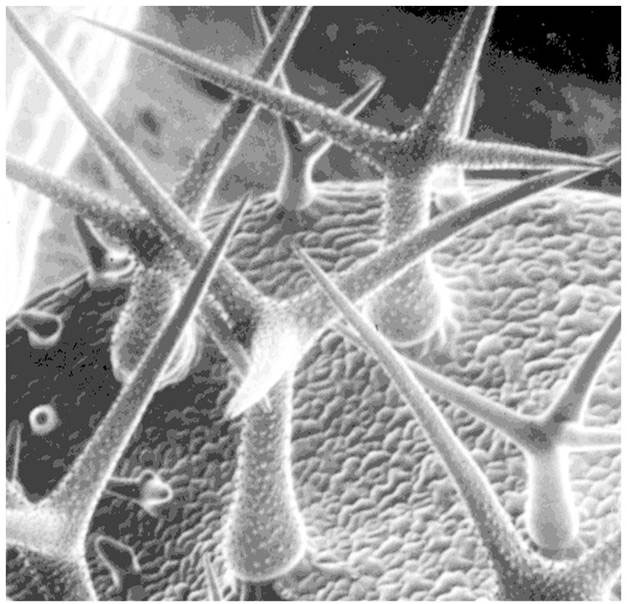
Покривните власинки по семената на памука, известни още като влакна на памука, са едноклетъчни, дълги до 7 cm, с клетъчни стени надебелени само от целулоза, което обуславя високото качество на памучните тъкани.
Жлезисти власинки
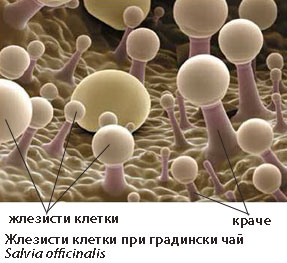 Основно имат защитна функция.
Основно имат защитна функция.- Жлезистите власинки синтезират различни химични съединения, които растенията използват за химична борба с бактерии, гъби, насекоми, тревопасни животни или за потискане на растежа на съседни растения от други видове.
- Изградени са от живи клетки (само от жлезисти клетки или само връхната клетка на власинката е жлезиста), които синтезират различни вещества (етерични масла, смолисти, слузести, парливи вещества) или отделят соли.
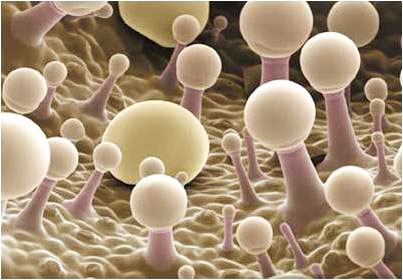
Подразделят се на:
- прости жлезисти власинки - могат да бъдат едноклетъчни или многоклетъчни. Многоклетъчните често имат "краче" и "главичка" → излъчваща жлезиста клетка на върха.
- сложни жлезисти власинки - те са многоклетъчни със сложен строеж, например тентаклите при насекомоядните растения.
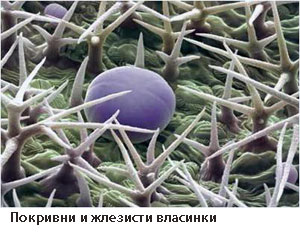 Органите на растенията не са покрити в еднаква степен с власинки
Органите на растенията не са покрити в еднаква степен с власинки
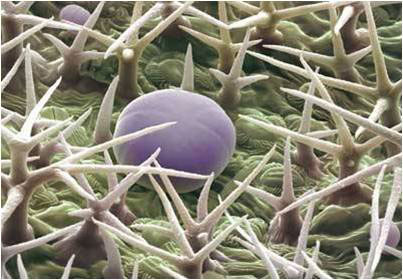
Те могат да са:
- голи - нямат власинки, или
- овласени - покрити с повече или по-малко власинки.
Според дължината на власинките и начина на ориентирането им, овласяването може да бъде:
- мъхесто овласяване - власинките са дълги, без ориентация;
- копринесто овласяване - власинките са тънки, дълги, ориентирани в една посока;
- паяжиновидно овласяване - власинките дълги, преплетени помежду си.
Kоркът замества епидермиса при наличие на вторично нарастване на растителните органи
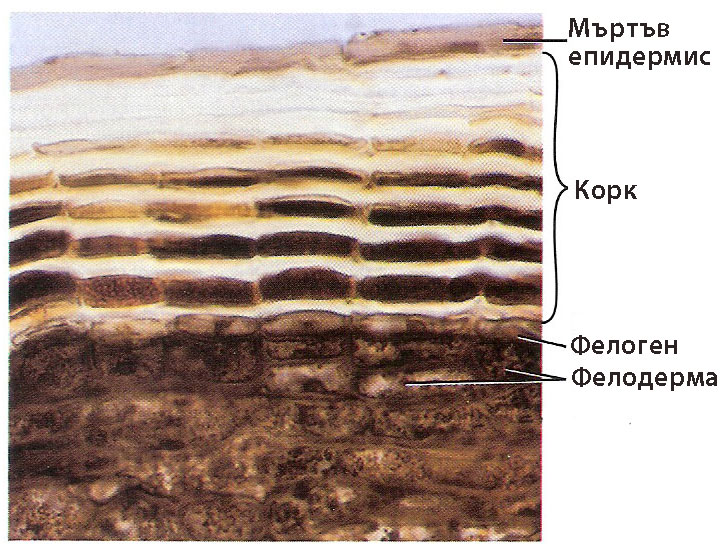 Коркът покрива растителните органи, които имат вторично нарастване. Развива се най-вече по стъблото и корените на дървесните видове.
Коркът покрива растителните органи, които имат вторично нарастване. Развива се най-вече по стъблото и корените на дървесните видове.
Корковата тъкан е изградена от няколко реда (понякога много)
- мъртви клетки,
- с правоъгълна форма и
- надебелени клетъчни стени пропити със суберин,
- разположени без междуклетъчни пространства.
Първоначално клетките на корка са живи. Клетъчните им стени са многослойни и еластични и постепенно суберинизират. Това довежда до отмиране на протопласта и напълно развитите коркови клетки са изпълнени с въздух.
Коркът е непропусклив за вода и газове и осигурява надеждна защита на растителните органи.
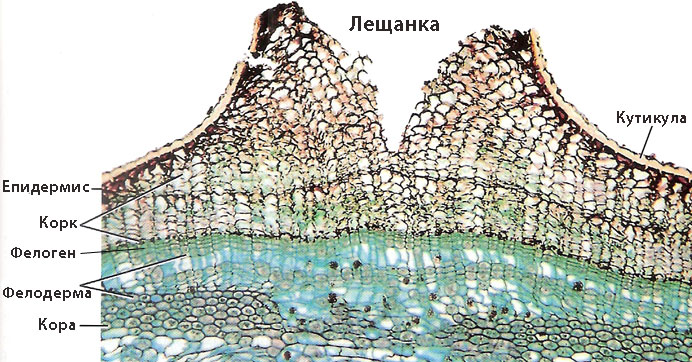
Аерирането на растителните органи покрити с корк се извършва през лещанки
Коркът е непропусклив за вода и газове и аерирането на растителните органи се извършва през специални образувания в корковата тъкан, наречени лещанки.
Лещанките се наблюдават като дребни, кръгли или продълговати брадавички или подутинки по кората на младите клони, и се виждат и с невъоръжено око. Те обикновено възникват под устицата и се образуват от собствен фелоген. Напълно развитите лещанки се състоят от рехаво разположени, закръглени, безцветни клетки с тънки несуберинизирани стени и значителни междуклетъчни пространства. В процеса на образуване на лещанките, епидермисът се разкъсва.
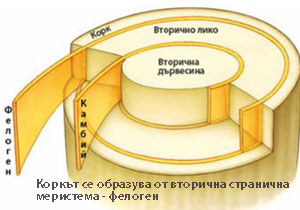 Коркът се образува от вторичната странична меристема фелоген или корков камбий
Коркът се образува от вторичната странична меристема фелоген или корков камбий
Първият фелоген се залага като цилиндър в осевите органи на растението. Най-често води началото си от паренхимните клетки разположени под епидермата, но при някои растения (зокум, круша) се образува от клетките на епидермата, а при други - от паренхимни клетки лежащи по-дълбоко в кората (лиственица, бор, касис).
Клетките на фелогена се делят тангенциално (успоредно на повърхността на растителния орган) и навън образуват корк, а навътре - фелодерма. Фелогенът образува много повече слоеве корк, отколкото фелодерма.
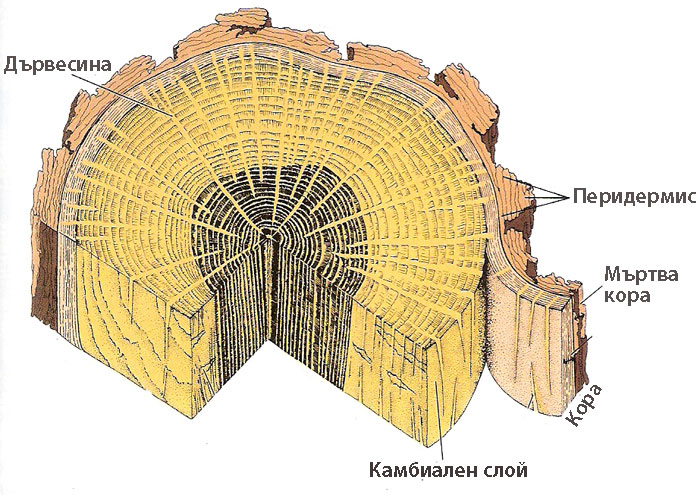
Трите слоя - корк, фелоген, фелодерма - са известни като перидермис.
Третична покривна тъкан - мъртва кора
Обикновено корковият камбий действа една година. Следващите години той се залага от по-дълбоко лежащи живи паренхимни клетки. Образуваният нов корк изолира не само живите тъкани на перидермиса, но и други тъкани на първичната кора разположени навън от него, които скоро умират. От ежегодното натрупване на тези мъртви тъкани се образува слой, наречен мъртва кора.
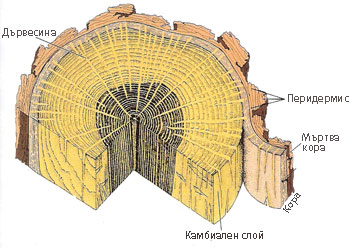 Мъртвата кора може да бъде неопадлива или опадлива
Мъртвата кора може да бъде неопадлива или опадлива
- Неопадлива - последователните слоеве на мъртвата кора са здраво свързани помежду си
- Опадлива - последователните слоеве на мъртвата кора не са здраво свързани помежду си
Неопадливата мъртва кора бива:
- гладка, или
- напукана
Опадливата мъртва кора бива:
- люсповидна (фелогенът се залага във вид на застъпващи се дъгички),
- пръстеновидна (фелогенът се залага като пръстен), и
- лентовидна
Проводящи тъкани
Проводящите тъкани при растенията провеждат водата и разтворените в нея минерални и органични вещества.
Биват два вида:
- Дървесинна проводяща тъкан (ксилема) - провежда водата и разтворените в нея минерални вещества от корена, през стъблото към листата, цветовете и плодовете
- Ликова проводяща тъкан (флоема) - провежда водата и разтворените в нея органични вещества в низходяща и възходяща посока - както от листата към всички останали органи, така и от корена към всички останали органи.
Ксилемата и флоемата заедно формират непрекъсната система, простираща се в цялото растение, наричана проводяща система.
Ксилемата и флоемата са комплексни тъкани, които се състоят от:
- проводящи елементи,
- паренхимни клетки, и
- склеренхимни влакна
 Дървесинна проводяща тъкан (ксилема)
Дървесинна проводяща тъкан (ксилема)
Ксилемата провежда водата и разтворените в нея минерални вещества от корена, през стъблото към листата
Ксилемата още има опорна функция и служи за отлагане на различни вещества.
Ксилемата е комплексна тъкан, която се състои от:
- Проводящи елементи: трахеиди и трахеи - провеждат водата и разтворените в нея минерални соли.
- Паренхимни клетки - живи клетки в които се натрупват различни съединения.
- Склеренхимни влакна - имат опорна функция, понякога в тях се отлагат различни вещества.
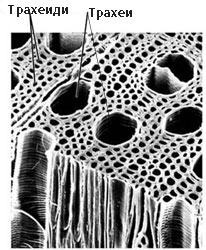
 Проводящи елементи на ксилемата - трахеиди и трахеи
Проводящи елементи на ксилемата - трахеиди и трахеи
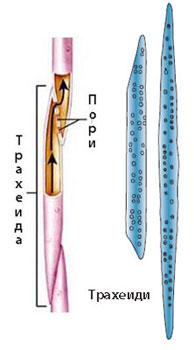
Трахеиди
Напълно развитите трахеиди са:
- Силно удължени мъртви клетки без протопласт, с надебелени и лигнифицирани клетъчни стени и заострени и силно скосени краища
- Дълги са средно 1 mm (4-7 mm при бор, 12 cm при лотос) и имат диаметър от няколко стотни до няколко десети от милиметъра
- Свързват се една с друга със скосените си краища
- Преминаването на водата от трахеида в трахеида става през пори
- Скоростта на водния ток през трахеидите е около 1m/h
Трахеидите са еволюционно по-стари и по-примитивни проводящи елементи в сравнение с трахеите. Те са единствените проводящи елементи за папратообразните и голосеменните растения
Трахеи (дървесинни цеви)
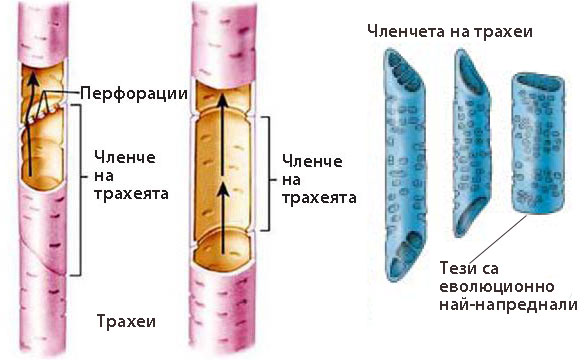 Напълно развитите трахеи представляват цеви (тръбички), изградени от множество лишени от протопласт клетки, наричани членчета на трахеите.
Напълно развитите трахеи представляват цеви (тръбички), изградени от множество лишени от протопласт клетки, наричани членчета на трахеите.
Членчетата на трахеите са:
- Повече или по-малко удължени мъртви клетки
- Клетъчните им стени са надебелени и лигнифицирани
- Напречните им стени са леко скосени или хоризонтални, перфорирани или напълно разрушени
В трахеите, членчетата на трахеите се свързват с напречните си стени и образуват цеви
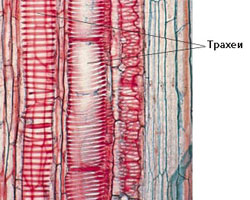 Напречните стени на членчетата на трахеите са перфорирани или напълно изчезнали;
Напречните стени на членчетата на трахеите са перфорирани или напълно изчезнали;- Това прави движението на водата безпрепятствено → съответно скоростта на движение е значително по-висока отколкото при трахеидите → достига 40-50 m/h;
- Дължината на трахеите средно е около 10 cm
(при някои растения достига до няколко метра → при дъба до 2 m, при лианите - до 4.5 m); - Диаметърът им е от 0.1 до 1 mm
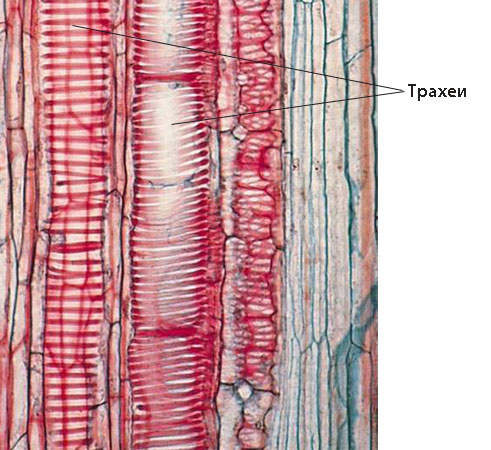
Перфорационни пластинки
- Понякога освен по напречните стени има перфорации и по страничните стени на членчетата на трахеите, което позволява движение на водата и в хоризонтална посока
- Тази част от стената на членчето на трахеята, която носи перфорацията се нарича перфорационна пластинка
- Перфорационните пластинки биват:
- прости - имат само една перфорация, и
- сложни - имат повече перфорации
- Сложните перфорационни пластинки са:
- стъпалчести - перфорациите са разположени успоредно една над друга подобно на стълба, и
- мрежести - перфорациите са разположени под формата на мрежа

Диференциране на трахеите
Диференцирането на трахеите е пример за програмирана смърт на клетките. Програмираната смърт на клетките е генетически програмиран процес, който при формирането на трахеите резултира в пълното разрушаване на протопласта. Клетъчните стени на членчетата на трахеите се запазват, с изключение на зоните на перфориране на напречните им стени. Диференцирането протича в следните стъпки:
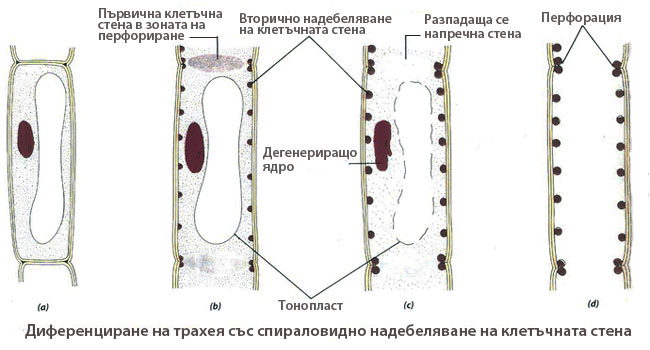
- (a) Младите членчетата на трахеите са живи, силно вакуолизирани клетки с първични клетъчни стени, свързани с напречните си стени.
- (b) Членчетата на трахеите нарастват предимно на широчина; започва спираловидно надебеляване на надлъжните им стени; напречните стени остават с първичен строеж и набъбват в зоната на перфорирането.
- (c) Завършва отлагането на вторичната клетъчна стена (спираловидното надебеляване в нашия случай), след което клетката преминава в състояние на автолиза. Ядрото дегенерира, тонопластът се разкъсва и напречните стени в зоните на перфорирането биват частично разрушени.
- (d) Членчето на трахеята вече е напълно диференцирано; протопластът му е напълно разрушен и двата му края са отворени в резултат на перфорирането на напречните му стени.
Трахеите са еволюционно по-млади и по-съвършени от трахеидите и произлизат от тях
Еволюцията на трахеите е вървяла към:
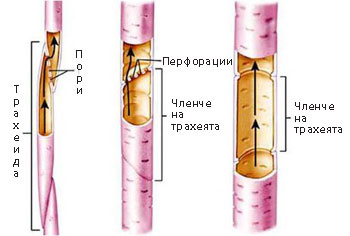 постепенното скъсяване и разширяване на отделните клетки-членчета
постепенното скъсяване и разширяване на отделните клетки-членчета- напречните стени на последните от коси и заострени стават хоризонтални
- перфориране или пълно изчезване на напречните стени на членчетата на трахеите
Широките трахеи осигуряват по-ефективен транспорт на водата и разтворените в нея вещества в сравнение с тесните трахеиди, но са склонни да се увреждат по-лесно.
Трахеите са основните проводящи елементи на покритосеменните растения, макар че ксилемата на много от цветните растения съдържа както трахеи така и трахеиди.
Паренхимни клетки на ксилемата
 Те са живи клетки, обикновено с лигнифицирани вторични клетъчни стени. По правило са разположени във вертикални ивици по дължината на проводящите елементи, а при наличие на вторично нарастване и в радиални лъчи.
Те са живи клетки, обикновено с лигнифицирани вторични клетъчни стени. По правило са разположени във вертикални ивици по дължината на проводящите елементи, а при наличие на вторично нарастване и в радиални лъчи.
Изпълняват различни функции:
→ участват в транспорта на веществата;
→ в тях се натрупват различни вещества (скорбяла, мазнини, танини, кристали и др.). Например, при листопадните дървета от умерената зона, в края на лятото в паренхимните клетки на ксилемата се отлага скорбяла, която през зимата се превръща в захароза и предпазва растителните органи от измръзване.
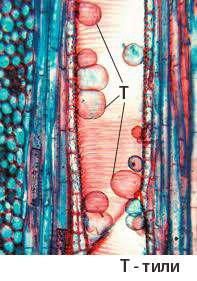
При някои растения паренхимните клетки на ксилемата образуват мехурести формирования, известни като тили, които навлизат в лумена на проводящите елементи и ги запушват
Образуването на тили се наблюдава предимно във вторичната ксилема на дървесните растения, в случаите когато трахеидите и трахеите:
- претърпят нараняване, последвано от загуба на вътрешното налягане;
- претърпят нападение от гъби и бактерии;
 застареят и престанат да участват в транспорта на веществата.
застареят и престанат да участват в транспорта на веществата.
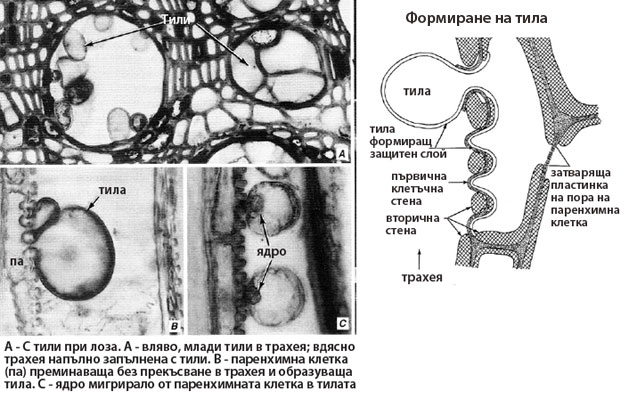
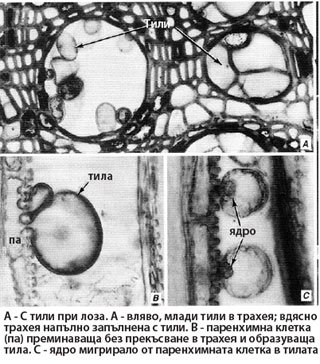
Тилите се образуват от паренхимни клетки на ксилемата, разположени в непосредствена близост до трахеите и трахеидите. Тези паренхимни клетки са наричани още контактни клетки, защото в буквалния смисъл на думата се намират в пряк контакт с проводящите елементи. Контактните клетки се характеризират с наличие на беден на целулоза и богат на пектинови вещества слой разположен върху вторичната им клетъчна стена, известен като защитен слой.
Формиране на тилите
- Затварящата пластинка на пората се разрушава по ензимен път.
- Защитният слой на контактната паренхимна клетка преминава през пората и образува мехуресто образувание (тила) в лумена на проводящия елемент.
- Ядрото на паренхимната клетка и част от цитоплазмата й също мигрират в тилата.
В тилите се натрупват различни вещества - най-често токсични фенолни съединения (танини, хинони, флавони), а някои тили могат да се диференцират и в склереиди. Тилите обикновено са многобройни и напълно запушват проводящите елементи. Така от една страна те усилват здравината на ксилемата, а от друга предотвратяват безпрепятственото развитие и разпространяване на патогенни бактерии и гъби в дървесината на растението.
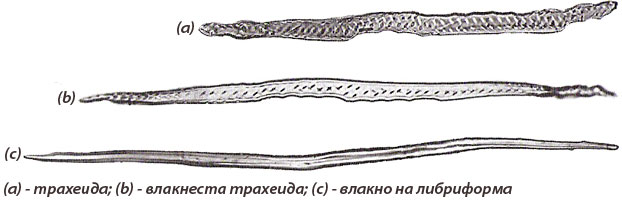
Склеренхимни влакна на ксилемата
Склеренхимните влакна придават здравина и устойчивост на ксилемата и биват:
- Влакнести трахеиди - по-слабо диференцирани склеренхимни влакна и по строеж заемат междинно положение между трахеидите и типичните склеренхимни влакна; те са по-дълги от трахеидите и дворчестите им пори са по-дребни.
- Влакна на либриформа - силно специализирани склеренхимни влакна, по-дълги от влакнестите трахеиди, с по-дебели клетъчни стени и прости пори по тях.
- Желатинови влакна - склеренхимни влакна, с надебелени клетъчни стени, притежаващи слоеве от хигроскопични вещества във вторичната си клетъчна стена. Тези вещества имат способност при определени условия да поглъщат вода и да набъбват.
Според произхода си ксилемата бива първична и вторична
- Първичната ксилема се образува от прокамбия
- Вторичната ксилема се образува от камбия
Първична ксилема бива протоксилема и метаксилема
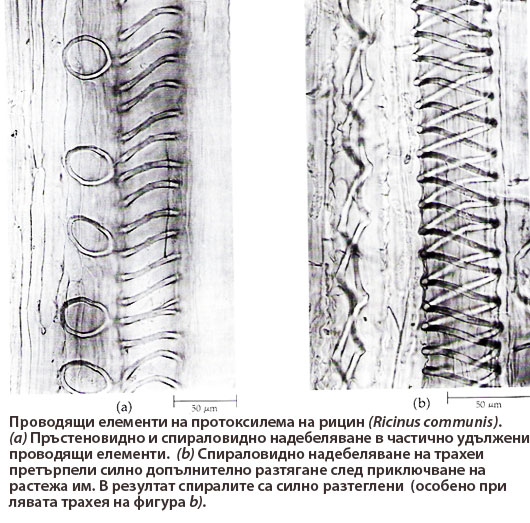
Протоксилема
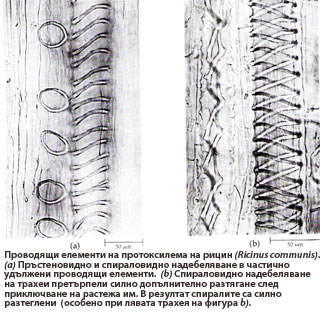 Формира се в периода на удължаване и нарастване на корена, стъблото и листата
Формира се в периода на удължаване и нарастване на корена, стъблото и листата- Тя е най-рано формираната проводяща тъкан (proto-, означава "първи")
- Обикновено проводящите й елементи имат пръстеновидно или спираловидно вторично надебеляване на клетъчните стени. Пръстеновидното и спираловидното надебеляване на клетъчните стени позволява допълнително разтягане на проводящите елементи след приключване на нарастването им.
- В протоксилемата видът на надебеляване на клетъчните стени на проводящите елементи силно се влияе от степента на удължаване на растителните органи → при значително удължаване се наблюдава пръстеновидно и спираловидно надебеляване на клетъчните стени; при слабо удължаване - стъпаловидно и мрежесто надебеляване.
- Проводящите елементи на протоксилемата обикновено се деформират и разрушават в процеса на удължаване и нарастване на растителните органи.
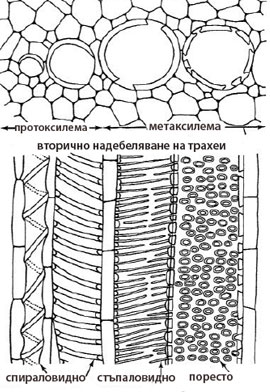 Метаксилема
Метаксилема
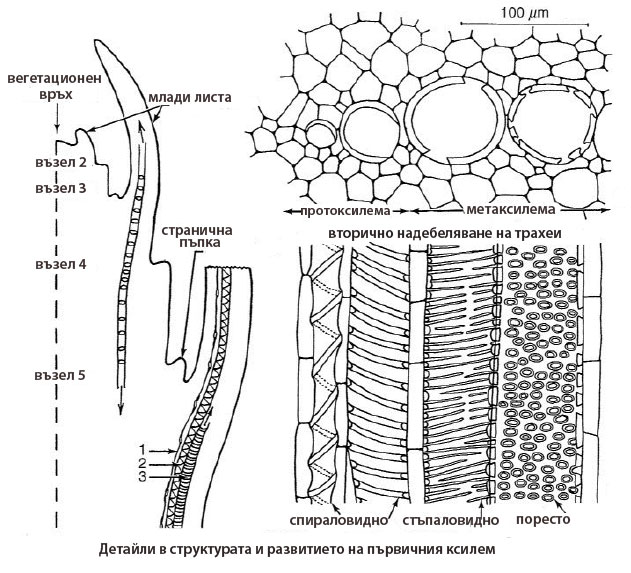
- Завършва развитието си след приключване удължаването на растителните органи.
- meta-, означава "след".
- Надебеляването на клетъчните стени основно е поресто.
- Клетъчните стени на проводящите елементи на метаксилемата са нееластични и твърди и не се поддават на допълнително разтягане.
- Проводящите елементи на метаксилемата са по-широки от тези на протоксилемата и не се разрушават.
Вторичната ксилема се образува от камбия
- Проводящите елементи на вторичната ксилема имат поресто надебеляване на клетъчните стени.
- Клетъчните стени на проводящите елементи са твърди и нееластични и не се поддават на допълнително разтягане
Ликова проводяща тъкан (лико, флоема)
Флоемата провежда разтворените във вода органични вещества в низходяща и възходяща посока
По ликовата проводяща тъкан освен захари се транспортират аминокиселини, липиди, хормони, голям брой различни протеини, някои от които служат като сигнални молекули, както и някои растителни вируси.
Флоемата е комплексна тъкан, която се състои от: 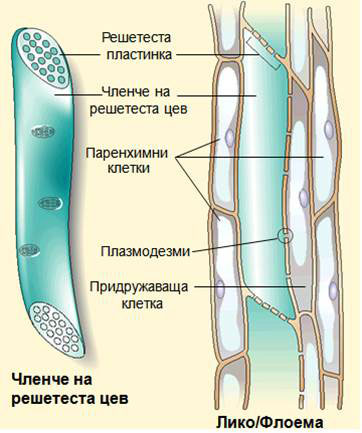
- Проводящи елементи: решетести клетки и решетести цеви с придружаващи клетки - по тях се движат разтворените във вода органични вещества.
- Паренхимни клетки - живи клетки в които се отлагат различни съединения.
- Склеренхимни влакна и склереиди - придават здравина и предпазват от деформиране на проводящите елементи.
Проводящи елементи на флоемата: решетести клетки и решетести цеви
Решетестите клетки са живи, дълги, тесни, със заострени краища и са характерни за голосеменните растения.
Решетестите цеви са характерни за покритосеменните растения.
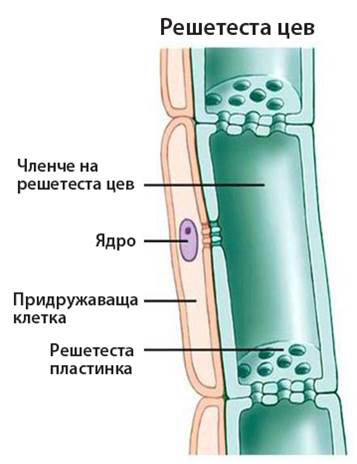
Те са изградени от отделни клетки, наричани членчета на решетестата цев. Последните имат хоризонтални напречни стени, по които се разполагат голям брой решетести полета (участъци от клетъчната стена пронизани от множество пори), което резултира във формиране на решетеста пластинка.
За разлика от членчетата на трахеите, напълно развитите членчета на решетестите цеви са живи. Обаче, в процеса на диференциране на решетестите цеви, протопластът на членчетата претърпява значителни промени, които водят до разграждане на:
- ядрото,
- рибозомите,
- апарата на Голджи,
- цитоскелета и
- тонопласта.
При окончателно развитите решетести цеви, протопластът е разположен до надлъжните стени на членчетата на решетестите цеви и от компонентите му са останали:
- добре развита гладка ендоплазмена мрежа,
- известно количество митохондри и пластиди, и
- специален Ф-протеин (Ф идва от Флоем)
Няма единно мнение за ролята на Ф-протеина, но някои ботаници смятат, че заедно с раневата калоза, той участва в запушването на решетестите полета при нараняване на решетестите цеви. В нормално състояние порите на решетестите полета са тапицирани с Ф-протеин, но той не ги запушва и не възпрепятства транспорта на веществата.
Така, при формиране на решетестите цеви се наблюдава частично разграждане на протопласта, за разлика от членчетата на трахеите, при диференцирането на които протича пълно разграждане на протопласта.
Придружаващите клетки са специализирани паренхимни клетки, които са тясно свързани с членчетата на решетестите цеви. Те притежават всички компоненти характерни за растителната клетка (включително и ядро). Членчетата на решетестите цеви и асоциираните с тях придружаващи клетки произлизат от една майчина клетка и имат множество плазмодезмени връзки помежду си. При липсата на ядро и рибозоми в протопласта на членчетата на решетестите цеви, придружаващите клетки са тези, които ги снабдяват с информационни молекули, протеини и АТФ, необходими за функционирането им.
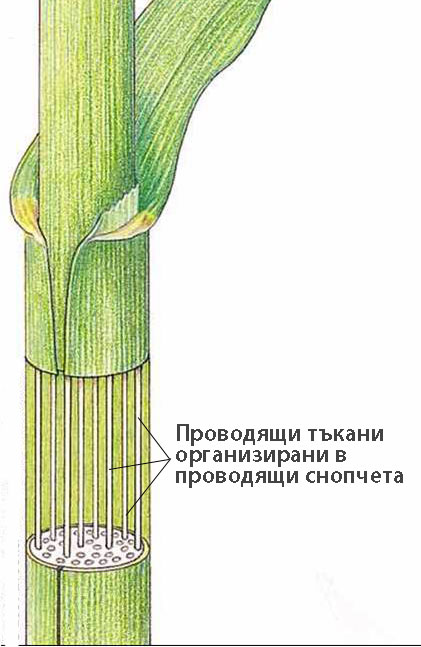
Проводящи снопчета
Проводящите тъкани са разположени заедно и образуват проводящи снопчета.
В зависимост от начина по който ксилемата и флоемата се разполагат една спрямо друга, проводящите снопчета биват колатерални, концентрични и радиални.
Колатерални проводящи снопчета
При тях ксилемата и флоемата се разполагат странично една спрямо друга. Колатералните проводящи снопчета се подразделят на:
- Отворени колатерални проводящи снопчета - между ксилемата и флоемата има камбий
- Затворени колатерални проводящи снопчета - между ксилемата и флоемата няма камбий
- Биколатерални проводящи снопчета - като отворените колатерални, плюс още един слой флоема от външната страна на ксилемата.
Концентрични проводящи снопчета
При тях едната проводяща тъкан обгражда другата. Те се подразделят на:
- Периксилемни проводящи снопчета - ксилемата обгражда флоемата
- Перифлоемни проводящи снопчета - флоемата обгражда ксилемата
Радиално проводящо снопче
При него ксилемата и флоемата се редуват. Наблюдава се в централния цилиндър на първично устроения корен.