Клетка
Всички организми са изградени от клетки
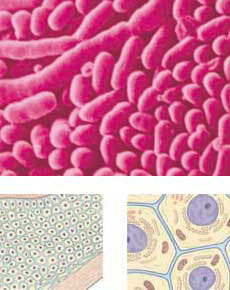 Някои организми се състоят от единични клетки, прекалено малки по размер за да бъдат видени с невъоръжено око, докато други са изградени от милиарди клетки. Ние не можем да си представим организъм, който да не е клетъчен по природа.
Някои организми се състоят от единични клетки, прекалено малки по размер за да бъдат видени с невъоръжено око, докато други са изградени от милиарди клетки. Ние не можем да си представим организъм, който да не е клетъчен по природа.
Клетката е структурна и функционална единица на всички организми и в нея протичат всички жизнени процеси.
Клетъчна теория
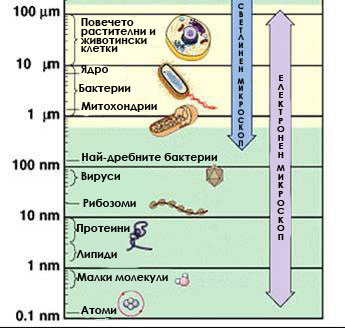
Основна характеристика на клетките е техният микроскопичен размер. С малки изключения типичната еукариотна клетка е с диаметър 10-100μm. Поради малкият им размер те са били открити едва след изобретяване на микроскопа в средата на 17-ти век. През 1665 година Роберт Хук, английски математик, физик и архитект, първи наблюдава и описва “мехурчета” и “празнини” в прерези на коркова тъкан и ги нарича “клетки”. Постепенно с усъвършенстването на микроскопа са открити ядрото, цитоплазмата, хлоропластите. Изяснява се, че организмовият свят е представен от едноклетъчни и многоклетъчни организми. Широкото разпространение на едноклетъчните организми затвърждава представата, че клетката е не само самостоятелна структурна, но и самостоятелна физиологична единица. Създават се предпоставки за формирането на обща научнообоснована теория за строежа на живите организми – клетъчната теория. Нейни създатели са Матиас Шлайден (ботаник) и Теодор Шван (зоолог) през 1838/1839 година.
Клетъчната теория в съвременната си форма включва следните три основни принципа:
1. Всички организми са съставени от една или много клетки.
2. Клетките са най-малката жива структурна и физиологична единица и всички процеси на метаболизма и наследствеността протичат в тях.
3. Клетките възникват само чрез делене на вече съществуващи клетки. Макар че живота вероятно е възникнал спонтанно, в днешно време биолозите не наблюдават възникването на нови клетки.
Клетката е оградена с мембрана единица, която съдържа ДНК и цитоплазма
 Когато се сравняват различни сложно устроени организми, на клетъчно ниво те се оказват забележително сходни. Например, макар че в човешкото тяло има над 100 различни типа клетки, те всички имат сходни черти и много от характеристиките им даже са сходни с тези на растенията.
Когато се сравняват различни сложно устроени организми, на клетъчно ниво те се оказват забележително сходни. Например, макар че в човешкото тяло има над 100 различни типа клетки, те всички имат сходни черти и много от характеристиките им даже са сходни с тези на растенията.Независимо, че строежа на клетките при различните организми варира, всички клетки притежават следните три основни структури:
- Плазмена мембрана
- Ядро или нуклеоид и
- Цитоплазма
Плазмената мембрана обгражда клетката
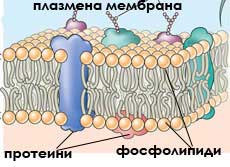 Плазмената мембрана обгръща клетката и изолира съдържанието й от външната среда. Тя представлява двоен фосфолипиден слой с дебелина от 5 до 10 nm, в който са разпръснати белтъчни молекули.
Плазмената мембрана обгръща клетката и изолира съдържанието й от външната среда. Тя представлява двоен фосфолипиден слой с дебелина от 5 до 10 nm, в който са разпръснати белтъчни молекули.
В напречен пререз, на електронен микроскоп плазмената мембрана се наблюдава, като две тъмни линии, разделени от по-светла зона. Тази картина се получава, поради специфичния начин на подреждане на фосфолипидните молекули – “опашка-към-опашка” и формиране на хидрофилни и хидрофобни зони в мембраната.
Протеините са вградени във фосфолипидния слой и в най-висока степен са отговорни за способността ѝ да взаимодейства с околната среда.
→ Транспортните протеини помагат на молекулите и йоните да се движат през плазмената мембрана навътре или навън от клетката.
→ Рецепторните протеини предизвикват промени в клетката, когато влязат в контакт с определени молекули, например хормони.
→ Маркерите идентифицират какъв тип е клетката, което е изключително важно при многоклетъчните организми, чиито клетки трябва да са способни да се разпознават, доколкото формират тъкани.
В централната част на клетката е разположен генетичният материал
 Всяка клетка съдържа поне една ДНК молекула, носеща наследствената информация.
Всяка клетка съдържа поне една ДНК молекула, носеща наследствената информация.
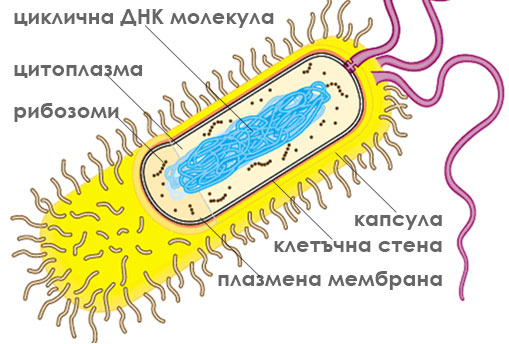
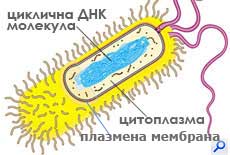
При прокариотите (бактериите), повечето от генетичният материал се намира в единствена циклична ДНК молекула, разположена в централната част на клетката. ДНК молекулата с прилежащата до нея цитоплазма се нарича нуклеоид. Tази зона не е отделена от останалата част на клетката с мембрана.
При еукариотите ДНК е разположена в ядро, отделено от останалата част на клетката с две мембрани. Всяка ДНК молекула е свързана и пакетирана с протеини, които подпомагат дейността й. В различни периоди от жизнения цикъл на клетката тези протеини обуславят ДНК молекулата да бъде по-слабо спирализирана (като хроматинови нишки) или по-силно спирализирана (като хромозоми с различна форма). 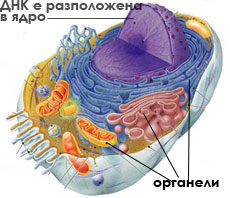 Както при прокариотите, така и при еукариотите, ДНК съдържа гените, които кодират синтеза на протеини в клетката и носят наследствената информация.
Както при прокариотите, така и при еукариотите, ДНК съдържа гените, които кодират синтеза на протеини в клетката и носят наследствената информация.
Цитоплазмата изпълва останалата част на клетката
Полутечен матрикс, наречен цитоплазма изпълва вътрешността на клетката (без ядрото/нуклеоида лежащо в нея). Цитоплазмата съдържа химичното богатство на клетката: захари, аминокиселини, протеини, мазнини, които клетката използва за да извършва ежедневната си дейност.
При еукариотните клетки, цитоплазмата притежава специализирани, ограничени с мембрани компартменти, наречени органели.
Типове клетъчна организация - прокариоти, еукариоти и мезокариоти
Прокариотните клетки са дребни, без сложна вътрешна организация.
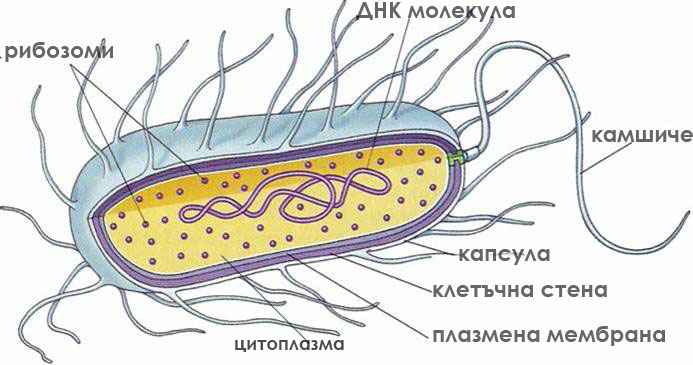 Прокариотите са разнообразни по форма, дребни клетки (0.1-3μm), с цитоплазма оградена от плазмена мембрана и здрава клетъчна стена. Някои от тях формират още капсула, реснички и камшичета. Те притежават повечето от метаболитичните процеси характерни за еукариотите, но тези процеси не протичат в мембранно ограничени органели. Вътрешната структура на прокариотните клетки е изключително просто организирана. Имат една циклична ДНК молекула разположена директно в цитоплазмата близо до централната част на клетката и рибозоми. Плазмената им мембрана образува вгъвания и извършва някои от функциите на липсващите органели. Размножават се с просто делене, без да се образува делително вретено. Възникнали са преди 3-3.5 милиарда години. Относително малък брой организми влизат в групата на прокариотите - микоплазми, бактерии и синьо-зелени делки.
Прокариотите са разнообразни по форма, дребни клетки (0.1-3μm), с цитоплазма оградена от плазмена мембрана и здрава клетъчна стена. Някои от тях формират още капсула, реснички и камшичета. Те притежават повечето от метаболитичните процеси характерни за еукариотите, но тези процеси не протичат в мембранно ограничени органели. Вътрешната структура на прокариотните клетки е изключително просто организирана. Имат една циклична ДНК молекула разположена директно в цитоплазмата близо до централната част на клетката и рибозоми. Плазмената им мембрана образува вгъвания и извършва някои от функциите на липсващите органели. Размножават се с просто делене, без да се образува делително вретено. Възникнали са преди 3-3.5 милиарда години. Относително малък брой организми влизат в групата на прокариотите - микоплазми, бактерии и синьо-зелени делки.
Еукариотните клетки имат сложна вътрешна организация
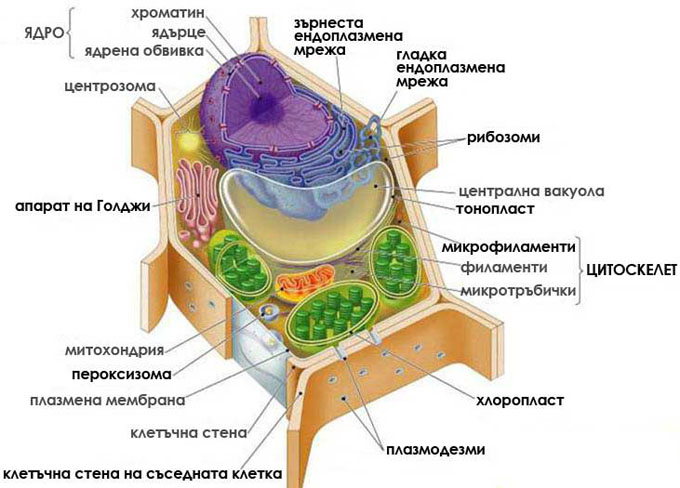 Еукариотните клетки имат далеч по-сложна вътрешна организация от прокариотите. Основна тяхна характеристика е компартменизацията. Те притежават голямо количество органели - мембранно ограничени структури, вътре в които едновременно и независимо едни от други протичат многобройни биохимични процеси. Например, растителните клетки имат голяма, ограничена с единична мембрана централна вакуола, в която се съхраняват протеини, захари, пигменти, отпадни продукти. И растителните и животинските клетки притежават силно развита ендомембранна система от цистерни и мехурчета - ендоплазнена мрежа, апарат на Голджи, транспортни мехурчета, микротелца. ДНК молекулите са разположени в оградено с две мембрани ядро, като всяка ДНК молекула е спирализирана и компактизирана с протеини. Вътрешната структура на еукариотните клетки се поддържа от цитоскелет. Размерът им варира между 10-100μm. Деленето им е чрез митоза и мейоза. Възникнали са от прокариотите преди около 1.5 милиарда години.
Еукариотните клетки имат далеч по-сложна вътрешна организация от прокариотите. Основна тяхна характеристика е компартменизацията. Те притежават голямо количество органели - мембранно ограничени структури, вътре в които едновременно и независимо едни от други протичат многобройни биохимични процеси. Например, растителните клетки имат голяма, ограничена с единична мембрана централна вакуола, в която се съхраняват протеини, захари, пигменти, отпадни продукти. И растителните и животинските клетки притежават силно развита ендомембранна система от цистерни и мехурчета - ендоплазнена мрежа, апарат на Голджи, транспортни мехурчета, микротелца. ДНК молекулите са разположени в оградено с две мембрани ядро, като всяка ДНК молекула е спирализирана и компактизирана с протеини. Вътрешната структура на еукариотните клетки се поддържа от цитоскелет. Размерът им варира между 10-100μm. Деленето им е чрез митоза и мейоза. Възникнали са от прокариотите преди около 1.5 милиарда години.
Всички организми с изключение на бактериите, микоплазмите и синьо-зелените делки са изградени от еукариотни клетки.
Мезокариотите имат междинен тип на клетъчна организация
Те имат ядро с двойна мембрана и обособени хромозоми (като еукариотите), но хромозомите им не съдържат хистонови белтъци (като прокариотите). Протичането на митозaта е различно от това на еукариотите.
Сходства между прокариотните, мезокариотните и еукариотните клетки
Независимо от различията, прокариотните, мезокариотните и еукариотните клетки имат редица сходства. И при трите типа клетки:
- Белтъчният синтез се извършва по обща схема с участието на рибозоми
- Репликацията и синтезът на ДНК се извършва по един и същи принцип
- Енергетичните процеси са сходни
- Плазмените мембрани имат избирателна пропускливост и способност за активен транспорт навън и навътре в клетката
- Могат самостоятелно да се възпроизвеждат
На практика, прокариотите притежават по-голяма част от метаболитичните функции на еукариотите, но те не протичат в добре обособени компартменти (зони) наречени органели.
Еукариотните клетки произлизат от прокариотните
Съществуват две теории обясняващи произхода на еукариотната клетка.
Теория за филиацията - според която органелите в еукариотната клетката са възникнали постепенно, в резултат на вгъването на плазмената мембрана и обособяването на специализирани органели изпълняващи различни функции - ядро, ендоплазмена мрежа, апарат на Голджи, митохондрии, пластиди.
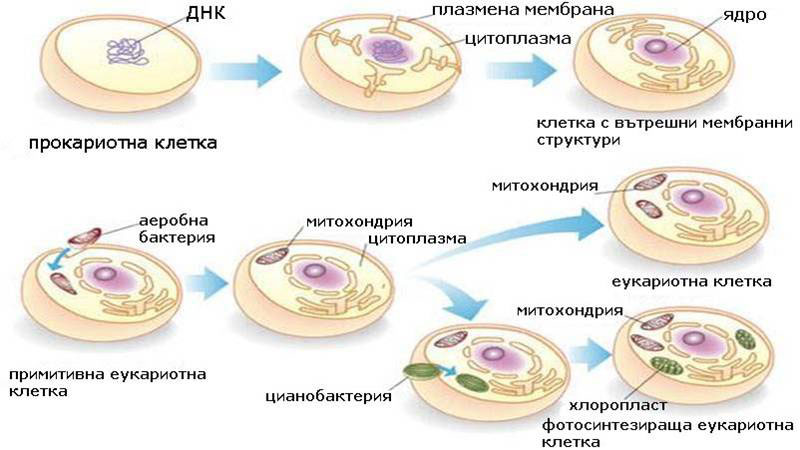 Теория за серийната ендосимбиоза - според която еукариотната клетка е произлязла по пътя на еволюцията, чрез обединяването на няколко типа (най-малко 4) самостоятелни прокариотни клетки и установяване на симбиотични взаимоотношения между тях. За родоначалник на еукариотните клетки се приемат анаеробни прокариоти (термоплазми) в които чрез фагоцитоза са проникнали: аеробни хетеротрофни бактерии, които са дали начало на митохондриите; спироплазми, които са внесли съкратителните белтъци; и цианобактерии, които са дали начало на хлоропластите.
Теория за серийната ендосимбиоза - според която еукариотната клетка е произлязла по пътя на еволюцията, чрез обединяването на няколко типа (най-малко 4) самостоятелни прокариотни клетки и установяване на симбиотични взаимоотношения между тях. За родоначалник на еукариотните клетки се приемат анаеробни прокариоти (термоплазми) в които чрез фагоцитоза са проникнали: аеробни хетеротрофни бактерии, които са дали начало на митохондриите; спироплазми, които са внесли съкратителните белтъци; и цианобактерии, които са дали начало на хлоропластите.
В подкрепа на теорията за ендосимбиозата има следните доказателства:
 1. Митохондриите и хлоропластите са сходни с бактериите по размер и устройство.
1. Митохондриите и хлоропластите са сходни с бактериите по размер и устройство.
2. Митохондриите и хлоропластите имат двойни мембрани с различен химичен строеж, като външната произлиза от плазмената мембрана на клетката домакин, а вътрешната е образувана от плазмената мембрана на аеробната бактерия.
3. И митохондриите и хлоропластите съдържат своя собствена циклична ДНК сходна с тази на бактериите.
4. Митохондриите и хлоропластите имат собствени (70S) рибозоми, с РНК сходна по състав с тези на бактериите (има същата последователност на аминокиселините)
5. Лабораторни изследвания показват, че амеби заразени от бактерии стават зависими от тях.
Защо клетките са малки?
 Повечето клетки са малки по практически причини, най-важната от които е комуникацията. Различните зони в клетката трябва да комуникират помежду си, за да може клетката да функционира ефективно. В клетката непрекъснато се извършва транспорт на химични вещества навътре и навън от нея. Съответно, колкото по-голям е размерът й, толкова повече време е необходимо на субстанциите да достигнат от плазмената мембрана до центъра на клетката и обратно. Поради тази причина организмите са изградени от много, малки по размер клетки, вместо от малко на брой, големи клетки. Предимството на малкия размер на клетките лесно се илюстрира чрез съотношението между повърхност и обем. Ако диаметъра на клетката се увеличава в порядък 1-2-3-4-5, повърхността й се увеличава 1-4-9-16-25, а обема - 1-8-27-64-125. Ясно се вижда, че увеличаването на обема на клетката не е последвано от еднакво увеличаване на повърхността на клетката. Всъщност, увеличаването на диаметъра 5 пъти, намалява повърхността на клетката към новия обем 5 пъти, в сравнение с изходния размер.
Повечето клетки са малки по практически причини, най-важната от които е комуникацията. Различните зони в клетката трябва да комуникират помежду си, за да може клетката да функционира ефективно. В клетката непрекъснато се извършва транспорт на химични вещества навътре и навън от нея. Съответно, колкото по-голям е размерът й, толкова повече време е необходимо на субстанциите да достигнат от плазмената мембрана до центъра на клетката и обратно. Поради тази причина организмите са изградени от много, малки по размер клетки, вместо от малко на брой, големи клетки. Предимството на малкия размер на клетките лесно се илюстрира чрез съотношението между повърхност и обем. Ако диаметъра на клетката се увеличава в порядък 1-2-3-4-5, повърхността й се увеличава 1-4-9-16-25, а обема - 1-8-27-64-125. Ясно се вижда, че увеличаването на обема на клетката не е последвано от еднакво увеличаване на повърхността на клетката. Всъщност, увеличаването на диаметъра 5 пъти, намалява повърхността на клетката към новия обем 5 пъти, в сравнение с изходния размер.
 Многоклетъчните организми обикновено се състоят от много на брой дребни клетки, понеже малките по размер клетки функционират по-ефективно. Те имат по-голяма относителна повърхност на единица обем и позволяват по-бърза комуникация между центъра на клетката и околната среда.
Многоклетъчните организми обикновено се състоят от много на брой дребни клетки, понеже малките по размер клетки функционират по-ефективно. Те имат по-голяма относителна повърхност на единица обем и позволяват по-бърза комуникация между центъра на клетката и околната среда.
Независимо, че клетките като правило са с малки размери (10-100μm), има клетки които са доста големи. Те са намерили начин да преодолеят проблема повърхност/обем, чрез един или повече адаптивни механизми. Някои от тях имат повече от едно ядра, други активно движат цитоплазмата си, а трети, като невроните например, са дълги и много тесни, така че във всеки даден момент цитоплазмата е близко до плазмената мембрана и по този начин транспорта на веществата остава бърз.
Различия между животинска и растителна клетка
Животинската клетка няма:
- Хлоропласти
- Вакуоли
- Клетъчна обвивка
- Плазмодезми
Растителната клетка няма:
- Камшичета - с изключение на някои сперматозоиди
Съатавни части на животинската клетка
Биологични мембрани
Взаимодействията на клетката с околната среда са сред най-важните дейности, които тя извършва и без тях живота й не може да съществува. Тези жизненоважни взаимодействия се извършват и контролират от плазмените (биологичните) мембрани.
Всяка плазмена мембрана е изградена от двоен фосфолипиден слой с потопени в него глобуларни протеини.
В този модел, наричан течно-мозаичен модел, фосфолипидните молекули формират полутечен двоен слой, а белтъчните молекули са мозаично разхвърляни в него.
Компоненти на клетъчната мембрана
Еукариотните клетки съдържат много, различни мембрани, които макар че не са идентични имат една и съща фундаментална структура. Всички клетъчни мембрани са изградени от следните четири компонента:
1. Двоен фосфолипиден слой
2. Трансмембранни протеини
3. Мрежа от поддържащи протеини
4. Повърхностни протеини и гликопротеини
1. Двоен фосфолипиден слой
Всяка клетъчна мембрана е изградена от фосфолипиден бислой, който осигурява пластичен матрикс за протеините и ограничава пропускливостта на мембраната.
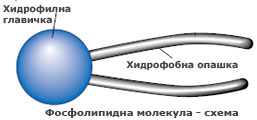
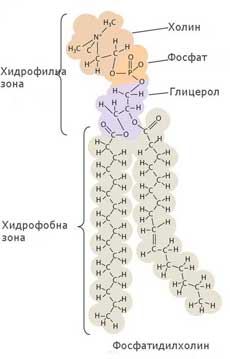 Фосфолипидите са сложни молекули, гръбнак на които е молекула глицерол с прикрепени към нея две мастни киселини (неполярни и съответно силно хидрофобни) и фосфорилиран алкохол (полярен и съответно силно хидрофилен). Понеже фосфорилираният алкохол обикновено е разположен от едната страна на молекулата, а двете въглеводородни вериги на мастните киселини от противоположната, фосфолипидите често се изобразяват като полярна главичка с две неполярни опашки.
Фосфолипидите са сложни молекули, гръбнак на които е молекула глицерол с прикрепени към нея две мастни киселини (неполярни и съответно силно хидрофобни) и фосфорилиран алкохол (полярен и съответно силно хидрофилен). Понеже фосфорилираният алкохол обикновено е разположен от едната страна на молекулата, а двете въглеводородни вериги на мастните киселини от противоположната, фосфолипидите често се изобразяват като полярна главичка с две неполярни опашки.
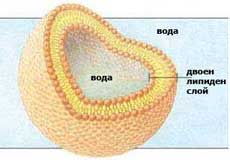
Когато фосфолипидни молекули бъдат поставени във вода, полярните молекули на водата отблъскват дългите неполярни опашки на фосфолипидите и те се оказват пакетирани плътно една до друга, сепарирани възможно най-далече от водата. Всяка фосфолипидна молекула се ориентира със своята полярна "главичка" навън към водата, а опашките винаги са ориентирани навътре и една към друга в бислоя. Те никога не контактуват с водата и формират неполярна бариера за водоразтворимите молекули. Фосфолипидните бислоеве възникват спонтанно и образуват или плоски слоеве или мембранни мехурчета
 Фосфолипидните двойни слоеве са течни с вискозитет на зехтин. Вискозитета им се променя с промяната на температурата, като се увеличава с намаляването й.
Фосфолипидните двойни слоеве са течни с вискозитет на зехтин. Вискозитета им се променя с промяната на температурата, като се увеличава с намаляването й.
Основно свойство на фосфолипидите е свободното им предвижване в бислоя.
2. Трансмембранни протеини
Главни компоненти на всяка мембрана е набор от протеини, потопени върху или във фосфолипидният двоен слой. Тези протеини осигуряват транспорт на вещества и информация през мембраната навътре и навън от клетката. Много от мембранните протеини не са фиксирани в постоянна позиция и могат да се движат в мембраната, както фосфолипидите. Някои мембрани имат голямо количество протеини, докато други ограничено.
3. Мрежа от поддържащи протеини
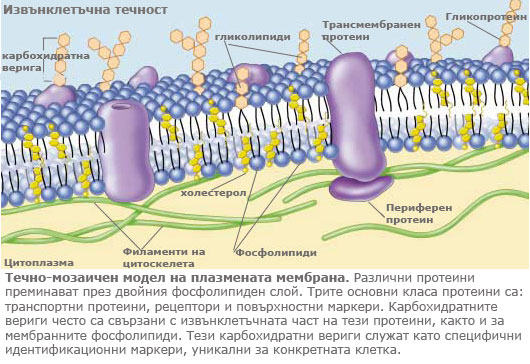 Мембраните са структурно подкрепяни от вътрешноклетъчни протеини, които определят формата на мембраната, като се свързват с актиновите филаменти на цитоскелета на клетката. Мембраните използват и мрежа от други протеини за да контролират страничното движение на някои ключови мембранни протеини, закотвяйки ги на определени места
Мембраните са структурно подкрепяни от вътрешноклетъчни протеини, които определят формата на мембраната, като се свързват с актиновите филаменти на цитоскелета на клетката. Мембраните използват и мрежа от други протеини за да контролират страничното движение на някои ключови мембранни протеини, закотвяйки ги на определени места
4. Повърхностни протеини и гликопротеини
Ендоплазмената мрежа синтезира и добавя вериги от захарни молекули на мембранните протеини и липиди, създавайки т.н. гликокаликс, който е разположен само от външната страна на мембраната. Различните типове клетки имат различни гликолипиди и гликопротеини по своята повърхност и те действат като маркери за идентичността на клетките.
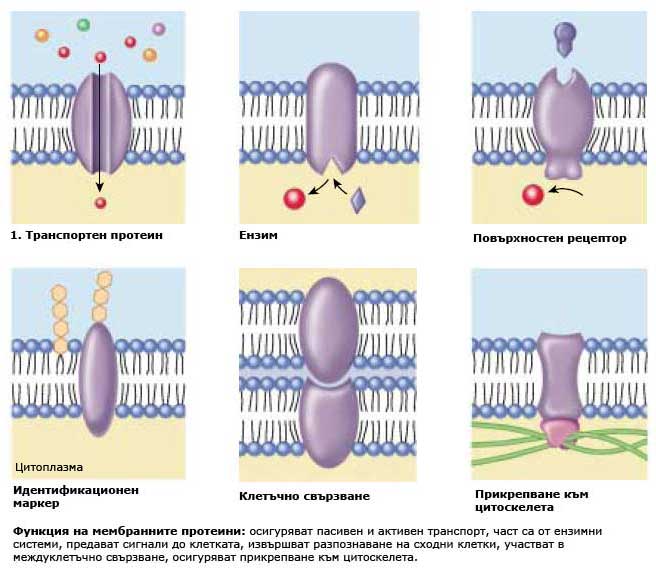
Функция на мембранните протеини
Независимо че клетките взаимодействат с околната среда по много различни начини, някои от тези взаимодействия директно включват мембранните протеини. Ние ще разгледаме шестте основни и ключови класа мембранни протеини и функциите които те изпълняват.
1. Транспортни протеини. Мембраните са силно избирателни, позволявайки само определени субстанции да влизат във или да напускат клетката. Транспорта на веществата се извършва през канали или чрез носители, по или срещу градиента на концентрация с участието на транспортните протеини.
2. Ензими. Клетките извършват много химични реакции върху вътрешната страна на плазмената мембрана, използвайки ензимите прикрепени към нея.
3. Рецепторни протеини. Мембраните са изключително чувствителни към химична информация, засичайки я чрез рецепторните си протеини, които действат като детектори.
4. Идентификационни маркери. Разположени са по външната повърхност на клетъчната мембрана и идентифицират клетката пред другите клетки. Повечето клетъчни типове притежават свои собствени комбинации от рецепторни протеини характерни само за съответния тип клетки.
5. Протеини за клетъчно свързване. Клетките използват специфични протеини за да се свързват една с друга. Някои от тези връзки са трайни, а други временни.
6. Протеини за прикрепване към цитоскелета. Протеините, които взаимодействат с другите клетки, често са закотвени към цитоскелета посредством прикрепящи протеини.
Ядро: информационният център на клетката
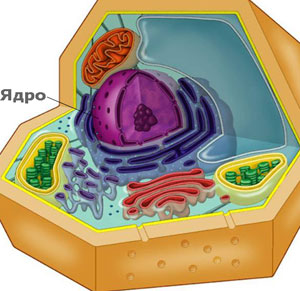 Ядрото е контролният център на клетката. То е най-голямата и най-лесно забележима органела на еукариотната клетка. В него е разполoжена генетичната информация, която командва цялата жизнена дейност в клетката. Ядрото отговаря за:
Ядрото е контролният център на клетката. То е най-голямата и най-лесно забележима органела на еукариотната клетка. В него е разполoжена генетичната информация, която командва цялата жизнена дейност в клетката. Ядрото отговаря за:
- функциониране и изграждане на клетката;
- реагиране на клетката на промените в околната среда;
- предаване на наследствената информация при делене на клетката;
- синтеза всички видове РНК, субединиците на рибозомите, ензими и др.
Повечето еукариотните клетки имат едно ядро, макар че клетките на някои гъби могат да имат от няколко до много ядра.
Форма на ядрото - обикновено сферична, но може да бъде елипсовидна, яйцевидна, пръчковидна, вретеновидна и дори звездовидна. Има корелация между формата на клетката и формата на ядрото.
Големина на ядрото – варира при различните растения и типове клетки. При клетките на висшите растения 4 -38 μm.
Месторазположение – непостоянно. В младите клетки винаги е в центъра. Като правило се предвижва в зони където има активно нарастване, нараняване или друга дейност.
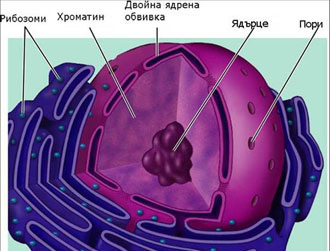 Строеж на ядрото
Строеж на ядрото
Ядрото има двойна обвивка от две биологични мембрани, затваряща течен матрикс в който са разположени хромозомите. В ядрото още се наблюдава тъмна зона наричана ядърце, където става интензивна синтеза на субединиците на рибозомите, на РНК и други протеини. Ядрената обвивка е покрита с голямо количество пори.
1. Ядрена обвивка
Ядрената обвивка е образувана от две клетъчни мембрани, с перинуклеарно пространство между тях от 20-40 nm. Външната мембрана на ядрената обвивка е свързана с цистерните на ендоплазмената мрежа в непрекъсната система. Някои автори дефинират ядрената обвивка като специализирана, локално диференцирана секция на ендоплазмената мрежа.
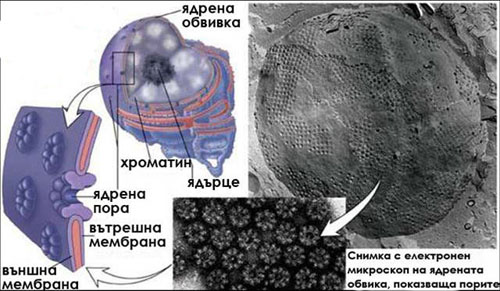 По повърхността на ядрената обвивка има множество ядрени пори. Те се формират на места където двете мембрани се прищипват заедно. Поровият канал е тапициран с протеини, които ограничават определени молекули да преминават във или навън от ядрото.
По повърхността на ядрената обвивка има множество ядрени пори. Те се формират на места където двете мембрани се прищипват заедно. Поровият канал е тапициран с протеини, които ограничават определени молекули да преминават във или навън от ядрото.
Транспорта е ограничен най-вече до:
(1) протеини влизащи в ядрото за да бъдат инкорпорирани в ядрените структури или да катализират химичните процеси протичащи в ядрото; и
(2) РНК и протеин-РНК комплекси формирани в ядрото и експортирани към цитоплазмата.
2. Ламина
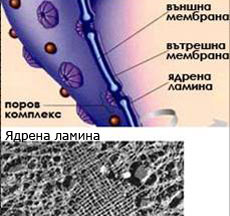
Представлява динамична нишковидна мрежеста структура от белтъчни фибрили, разположена непосредствено под ядрената обвивка. Ламината взема активно участие в дезинтегрирането и повторното изграждане на ядрената обвивка при деленето на клетката. Още участва в поддържането формата на ядрото.
3. Хромозомите: пакетиране на ДНК
И при еукариотите и при бактериите ДНК съдържа наследствената информация на клетката, която определя структурата и функцията на клетката. Но за разлика от цикличната ДНК (няма свободни краища) на прокариотите, ДНК на еукариотите е разделена на различен брой линейни хромозоми (имат два свободни края).
 С изключение на времето когато клетката се дели, нейните хромозоми са деспирализирани и разтегнати в тънки нишки наречени хроматинови нишки (представляват ДНК асоциирана с протеини, най-често в съотношение 40% ДНК и 60% протеини). Тази отворена нишковидна организация позволява протеините да се прикрепят към специфичната нуклеотидна последователност по дължината на ДНК молекулата. Без този достъп ДНК няма да е в състояние да контролира ежедневната дейност на клетката.
С изключение на времето когато клетката се дели, нейните хромозоми са деспирализирани и разтегнати в тънки нишки наречени хроматинови нишки (представляват ДНК асоциирана с протеини, най-често в съотношение 40% ДНК и 60% протеини). Тази отворена нишковидна организация позволява протеините да се прикрепят към специфичната нуклеотидна последователност по дължината на ДНК молекулата. Без този достъп ДНК няма да е в състояние да контролира ежедневната дейност на клетката.
Когато клетката се подготвя за делене, ДНК започва да се спирализира и кондензира. В началните етапи на тази спирализация групи от хистонови белтъци могат да бъдат видяни омотани с ДНК молекулата. Тези структури се наричат нуклеозоми и наподобяват на нанизани на нишка мъниста. Спирализирането продължава, докато ДНК премине в компактна маса. Тези напълно кондензирани хромозоми лесно се виждат под светлинен микроскоп в делящата се клетка. След разделянето на клетката хромозомите се деспирализират и не могат вече да се наблюдават със светлинен микроскоп. Деспирализирането на ДНК в хроматинови нишки позволява на ензимите да правят РНК копия на ДНК молекулата. Посредством тези РНК копия, информацията в ДНК се използва за синтеза на протеини.
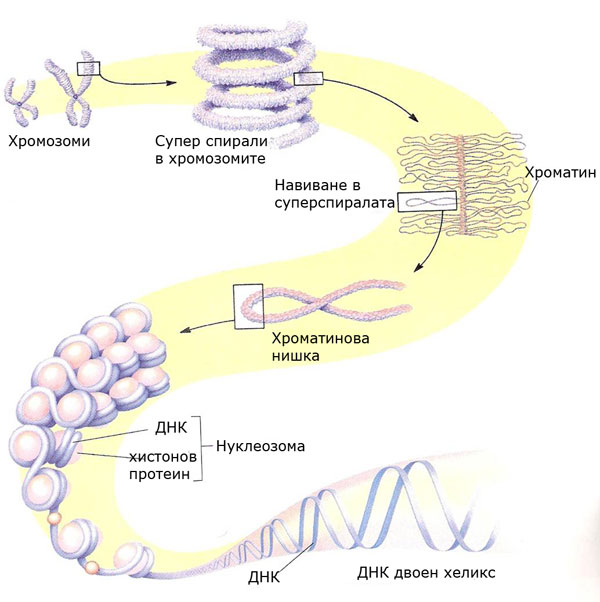 Хромозоми - структура
Хромозоми - структура
ДНК молекулата е изградена от две дълги нуклеотидни вериги, които имат нужда от допълнително пакетиране за да се съберат в ядрото.
ДНК молекулата плътно се навива около хистонови белтъци и образува нуклеозоми. Нуклеозома е група от 8 хистонови белтъчни молекули, всяка от които е обхваната от две завивания на ДНК молекулата.
Нуклеозомите продължават да се спирализират и компактизират и образуват хроматинови нишки.
Хроматиновите нишки продължават да се кондензират, като формират супер спирали и достигат форма известна като хромозома.
Морфология на метафазните хромозоми
Всяка метафазна хромозома има центромера – стеснен участък в хромозомата, който (1) прищъпва хромозомата и я разделя на две рамена и (2) участва в прикрепването на хромозомите за нишките на делителното вретено.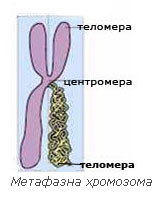 Според мястото на центромерата и дължината на рамената на хромозомите те биват:
Според мястото на центромерата и дължината на рамената на хромозомите те биват:
- Метацентрични хромозоми – имат приблизително равни рамена
- Субметацентрични хромозоми – рамената не са равни
- Акроцентрични хромозоми – имат едно много дълго и едно много късо рамо
Теломерите са зони в двата края на всяка хромозома. Те са изключително важни за живота на клетката. Теломерите предпазват краищата на различните хромозоми от случайно свързване и заедно с теломеразата определят продължителността на живота на клетките.
Хромозомен набор
 Броят, размерите и морфологичните особености на хромозомите са постоянни и характерни и за всеки вид и образуват неговия кариотип. Броят (наборът) на хромозомите, които организмите съдържат в телесните (соматичните) си клетки варира много силно между различните видове. Лукът (Allium cepa) има 16 хромозоми в клетка, зелето (Brassica oleracea) - 18 , пшеницата (Triticum vulgare) - 42; докато някои папрати имат над 1000 хромозома (Othioglossum spp. има около 1250).
Броят, размерите и морфологичните особености на хромозомите са постоянни и характерни и за всеки вид и образуват неговия кариотип. Броят (наборът) на хромозомите, които организмите съдържат в телесните (соматичните) си клетки варира много силно между различните видове. Лукът (Allium cepa) има 16 хромозоми в клетка, зелето (Brassica oleracea) - 18 , пшеницата (Triticum vulgare) - 42; докато някои папрати имат над 1000 хромозома (Othioglossum spp. има около 1250).
Хромозомния брой бива два вида - диплоиден и хаплоиден
- Диплоидният хромозомен брой ("двоен набор от хромозоми") е характерен е за телесните клетки. Той се получава след оплождане и се обозначава с 2n.
- Хаплоидният хромозомен брой ("единичен набор от хромозоми") е характерен за половите клетки и спорите. Получава се в резултат на редукционно делене, обозначава се с n и е равен на половината от диплоидния хромозомен брой.
Клетки, които имат повече от два набора хромозоми се наричат полиплоидни (3n, 4n, 5n, или повече).
При оплождането се възстановява диплоидния хромозомен брой, характерен за вида. Диплоидният хромозомен набор е представен от двойки хомоложни хромозоми - еднакви по форма и големина хромозоми, едната идваща от мъжката, а другата - от женската гамета при оплождането.
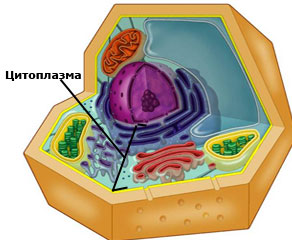 Цитоплазма – съдържанието на клетката затворено между плазмената мембрана и ядрото.
Цитоплазма – съдържанието на клетката затворено между плазмената мембрана и ядрото.
Състои се от:
- Цитозол / хиалоплазма - течната част на цитоплазмата, включваща:
- течен матрикс - фракция, с голям брой водоразтворими органични и неорганични вещества и разположени в него:
- рибозоми
- микротелца: пероксизоми, глиоксизоми, транспортни мехурчета
- микрофиламенти, филаменти и микротръбички - Ендоплазмена мрежа (ЕМ),
- Диктиозоми
- Митохондрии
- Пластиди
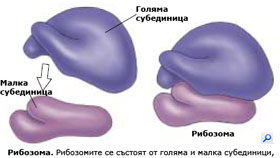 Рибозоми: място на протеиновия синтез в клетката
Рибозоми: място на протеиновия синтез в клетката
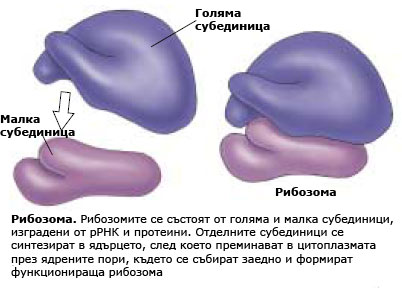
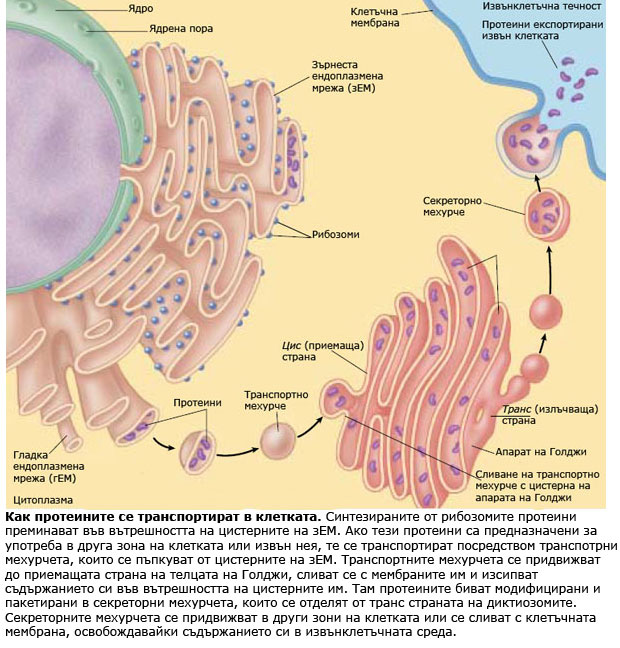
Независимо че ДНК в клетъчното ядро кодира последователността на аминокиселините в протеините на клетката, протеините не се синтезират в ядрото, а в цитоплазмата. Синтеза на протеините протича в РНК-протеинови комплекси наречени рибозоми.
Рибозомите са малки телца, изградени от рибозомна РНК свързана с комплекс от протеини в съотношение около 50/50. Рибозомите са сред най-комплексните структури на клетката. Всяка рибозома е изградена от две субединици (голяма и малка субединица), които се събират заедно 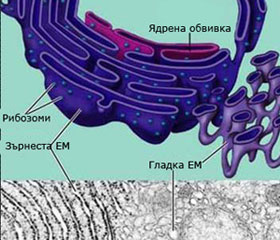 и образуват функционираща рибозома, само в момента на синтез на протеини. Субединиците на рибозомите се синтезират в ядърцето, но функционират в цитоплазмата. Някои рибозоми са разположени свободно в цитозола, други са прикрепени към ендоплазмената мрежа (ЕМ) и ядрената обвивка. Протеините, които функционират в цитоплазмата са синтезирани от свободните рибозоми разположени там, докато протеините включени в мембраните или предназначени за експорт извън клетката се произвеждат от рибозомите прикрепени към ЕМ.
и образуват функционираща рибозома, само в момента на синтез на протеини. Субединиците на рибозомите се синтезират в ядърцето, но функционират в цитоплазмата. Някои рибозоми са разположени свободно в цитозола, други са прикрепени към ендоплазмената мрежа (ЕМ) и ядрената обвивка. Протеините, които функционират в цитоплазмата са синтезирани от свободните рибозоми разположени там, докато протеините включени в мембраните или предназначени за експорт извън клетката се произвеждат от рибозомите прикрепени към ЕМ.
Броят на рибозомите в клетката зависи от метаболитичната й активност и варира от няколко стотин хиляди до няколко милиона. Рибозомите активно участващи в протеиновия синтез обикновено са разположени на групи или агрегати, наричани полизоми или полирибозоми.
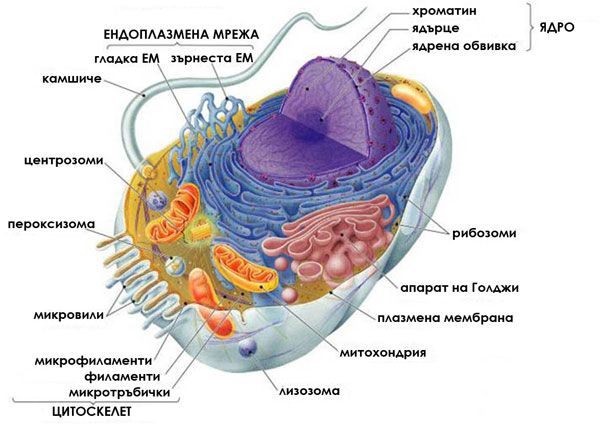
Ендоплазмена мрежа (ЕМ): компартменизира клетката
Ендоплазмената мрежа изпълва вътрешността на еукариотната клетка. Представлява обширна система от взаимно свързани, силно разклонени цистерни и канали, започващи от ядрото и достигащи до плазмалемата. Цистерните и каналите са изградени от плазмени мембрани без свободни краища и с около 4 nm лумен между тях. Тези цистерни и канали сепарират вътрешността на клетката на зони, канализирайки движението на молекулите в тях и осигурявайки повърхност за синтез на липиди и някои протеини. ЕМ е продължение на външната мембрана на ядрото.
ЕМ бива два типа:
- зърнеста ЕМ - с прикрепени рибозоми по нея
- гладка ЕМ – няма прикрепени рибозоми по нея
Видът и степента на развитие на ЕМ варира силно от клетка до клетка и зависи от типа на клетката, метаболитичната й активност и стадия и на развитие. Ендоплазмените мембрани са в непрекъсната динамика и постоянно променят своята форма и разположение.
Зърнеста ЕМ
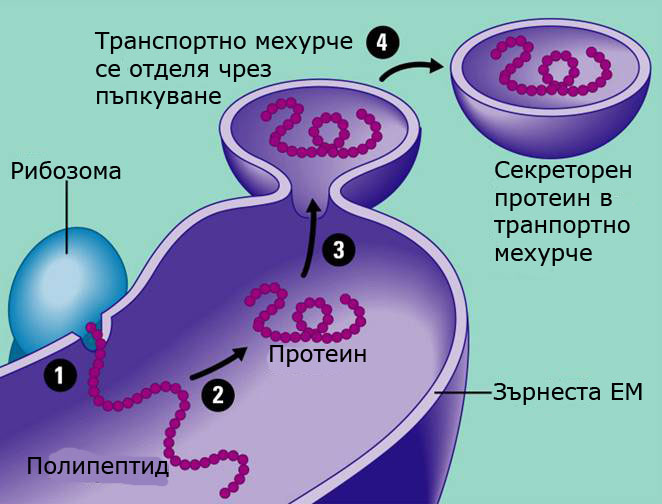 Изградена е основно от цистерни и главната й функция е синтеза и съхраняване на протеини, които ще бъдат експортирани навън от клетката. В районите където се извършва синтеза на протеини мембраните на ЕМ са обилно покрити с рибозоми и изглеждат зърнести.
Изградена е основно от цистерни и главната й функция е синтеза и съхраняване на протеини, които ще бъдат експортирани навън от клетката. В районите където се извършва синтеза на протеини мембраните на ЕМ са обилно покрити с рибозоми и изглеждат зърнести.
Синтезираните от рибозомите протеини преминават през мембраната във вътрешността на цистерните и от там чрез транспортни мехурчета се предвижват до вътрешната повърхност на плазмалемата, откъдето се освобождават навън от клетката.
Гладка ЕМ
Има тубуларна структура с малко количество рибозоми. Мембраните й съдържат множество ензими, които катализират синтеза на различни карбохидрати и липиди (фосфолипиди, масла, стероиди, секс хормони). Клетките, които извършват интензивна липидна синтеза, имат силно развита гладка ЕМ - например клетките на черния дроб, мозъка и пр. В черния дроб ензими от гладката ЕМ участват в детоксикацията на лекарства и отрови, а също и в обмяната на въглехидратите - в превръщането на гликогена до глюкоза.
Синтезираните липиди от гладката ЕМ се освобождават в цитозола като мастни капки. Те са повече или по-малко сферични структури, неоградени с плазмена мембрана и се наричат сферозоми. Сферозомите са особено изобилни в клетките на плодовете и семената. Около 45% от теглото на семената на слънчогледа, сусама, фъстъка, лена се дължи на съдържащите се в тях масла. Липидите осигуряват енергия и са източник на въглерод за поникващите растения.
ЕМ на съседните растителни клетки е свързана посредством цитоплазмени нишки, наричани плазмодезми. Те пресичат клетъчните стени и играят важна роля в комуникацията на клетките.
Апарат на Голджи: транспортната система на клетката
 Апаратът на Голджи е транспортната система на еукариотната клетка. Той получава, видоизменя, кондензира, пакетира и транспортира различни химични съединения синтезирани от ендоплазмената мрежа и подлежащи на употреба в други зони на клетката.
Апаратът на Голджи е транспортната система на еукариотната клетка. Той получава, видоизменя, кондензира, пакетира и транспортира различни химични съединения синтезирани от ендоплазмената мрежа и подлежащи на употреба в други зони на клетката.
Телцата на Голджи (наричани още диктиозоми) са с диаметър около 2 μm и дебелина около 0.5 μm. Те се състоят от 5 - 20 разположени една над друга, плоски, леко извити, несвързани помежду си цистерни, които често се разклоняват в сложна система от тръбички по периферията си. Съвкупността от всички телца на Голджи се нарича апарат на Голджи. Броят на телцата на Голджи варира от 20 и повече в животинските клетки дo няколкостотин в растителните. Особено многобройни са те в клетките на жлезистите власинки на растенията, които синтезират и излъчват различни субстанции.
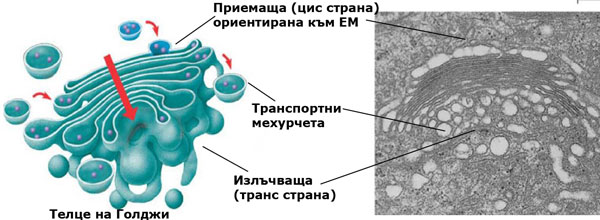
Апаратът на Голджи е динамична и силно поляризирана система. Телцата на Голджи имат предна или приемаща страна (цис страна) и задна или излъчваща страна (транс страна). Приемащата страна обикновено е ориентирана към ЕМ. Протеините и липидите синтезирани от ЕМ се транспортират до телцата на Голджи, и се модифицират като преминават в стъпаловиден порядък през цистерните. Най-честите трансформации са добавяне към молекулите им на захарни и липидни вериги и образуването на гликопротеини и гликолипиди. Новополучените или променени гликолипиди и гликопротеини се събират в краищата на цистерните и се отделят опаковани като секреторни мехурчета. Тези мехурчета след това се предвижват в съответните направления.
В растенията повечето от телцата на Голджи са ангажирани в синтеза, транспортирането и секретирането на гликопротеините и нецелулозните полизахариди предназначени за изграждането на клетъчната стена. Отделените мехурчета от телцата на Голджи достигнат плазмената мембрана и се сливат с нея, освобождавайки (секретирайки) съдържанието си в клетъчната стена.
Как протеините се транспортират и модифицират в клетката
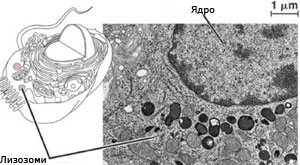
Лизозоми и микротелца
Лизозомите и микротелцата са мехурчета, които съдържат различни ензими. Изолирането на тези ензими в мехурчета защитава останалата част на клетката от нежелана химична активност.
Лизозоми: вътреклетъчните смилателни центрове
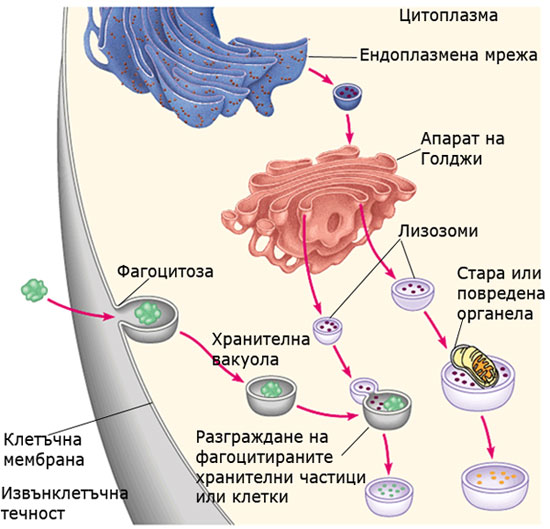 Лизозомите представляват мехурчета, отделяни от апарата на Голджи. Те съдържат хидролитични ензими (над 40 на брой), които катализират бързото разграждане на протеини, нуклеинови киселини, полизахариди, липиди и пр. Представляват “стомахa” на клетката и извършват вътрешно клетъчното разграждане на веществата. Хидролитичните ензими на лизозомите функционират в силно кисела среда.
Лизозомите представляват мехурчета, отделяни от апарата на Голджи. Те съдържат хидролитични ензими (над 40 на брой), които катализират бързото разграждане на протеини, нуклеинови киселини, полизахариди, липиди и пр. Представляват “стомахa” на клетката и извършват вътрешно клетъчното разграждане на веществата. Хидролитичните ензими на лизозомите функционират в силно кисела среда.
Лизозомите още:
- разграждат старите органели, като рециклират техните молекули и освобождават място за новообразуваните. Например митохондриите в клетките на някои тъкани биват заменяни с нови на всеки десет дни.
- участват в елиминирането на други клетки, които клетката е погълнала чрез фагоцитоза. Например, белите кръвни клетки използват лизозоми за разрушаване на фагоцитирани от тях бактериалните клетки.
- участват в т.н. "програмирана смърт на клетките” - процес при който протопласта на клетки, генетично програмирани да умрат (например склеренхимни и коркови клетки при растенията), бива разрушаван отвътре от техните лизозоми.
Механизъм на действие: лизозомите се сливат с транспортните мехурчета, хранителните вакуоли и всички останали мембранно заградени отпадъци. В новополученото мехурче освобождават ензимите си и разграждат намиращите се вътре материали до съставните им мономери.
Микротелца
Еукариотната клетка съдържа множество ензими-съдържащи мехурчета, ограничени от единични мембрани, наричани микротелца. Транспортирането на ензимите в микротелца е един от принципните начини, по който еукариотните клетки организират своя метаболизъм. 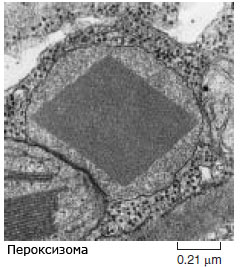 Микротелцата са с диаметър около 1-2 μm.
Микротелцата са с диаметър около 1-2 μm.
Към микротелцата на растителните клетки спадат:
- пероксизоми - сферични микротелца, с диаметър 0.5-1.5 μm. Имат грануларна вътрешна структура и могат да съдържат централна кристална структура с форма на диамант, изградена от протеин. Нямат вътрешни мембрани. В листата на растенията са тясно свързани с работата на хлоропластите и митохондриите и играят важна роля при фотодишането. Притежават способността да разграждат водородния прекис (оттам и името пероксизоми), отделящ се като страничен продукт от дейността на клетката. Водородният прекис е опасен за клетката поради високата си реактивоспособност.
- глиоксизоми - съдържат ензими, които трансформират мазнините до захароза по време на покълването на много семена.
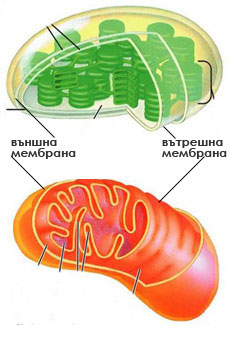
Митохондрии: източници на химична енергия за клетката
Митохондриите са дребни, безцветни органели с диаметър 0.5-1.5 μm и дължина 3-10μm. Те са открити във всички еукариотни клетки и броят им варира от една до 1000 в клетка.  Митохондриите имат две обвивки:
Митохондриите имат две обвивки:
- външна - гладка, и
- вътрешна - силно нагъната и формираща многобройни вгъвания, известни като кристи.
Кристите разделят митохондрията на две зони:
- вътрешна, лежаща навътре от вътрешната мембрана, и
- перимитохондриално пространство, разположено между двете мембрани.
На повърхността на вътрешната мембрана, а също и вградени в нея се намират протеините извършващи клетъчното дишане.
Вътрешността на митохондрията е изпълнена с течен матрикс, в който има множество ензими и митохондриални рибозоми, РНК и ДНК. Митохондриите имат своя собствена ДНК, контролираща синтеза на няколко протеина (но не на всички митохондриални протеини), ключови за клетъчното дишане. Синтеза на тези протеини става в матрикса на митохондриите с участието на митохондриални рибозомни комплекси.
Митохондриите могат да се сливат, да се делят и се намират в непрекъснато движение в клетката, като се натрупват в места където е необходима енергия. В клетки където клетъчната мембрана активно транспортира вещества, митохондриите могат да бъдат наблюдавани подредени непосредствено до нея.
Митохондриите увеличават броя си чрез просто делене. Контролът над деленето им се извършва от ядрото на клетката.
Пластиди
Пластидите, заедно с вакуолите и клетъчната стена, са характерни само за растителната клетка. Те са двойномембранни органели и според оцветяването си биват:
- Хлоропласти - зелени;
- Лейкопласти - безцветни;
- Хромопласти - жълти, червени, оранжеви
Трите вида пластиди са генетически родствени, но имат различна структура и изпълняват различни функции.
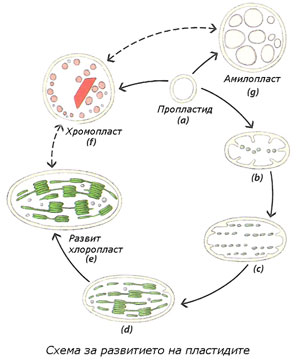 Пропластидите са предшественици и на трите вида пластиди
Пропластидите са предшественици и на трите вида пластиди
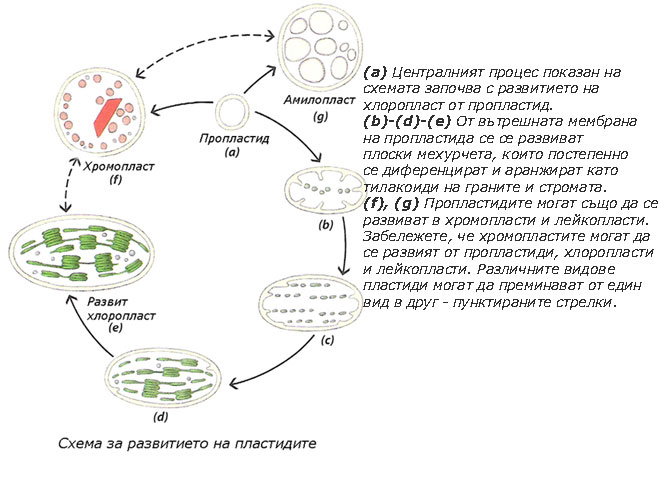
Пропластидите са малки, двумембранни, безцветни, недиференцирани пластиди, със слабо развита вътрешномембранна структура. Те се срещат в яйцеклетката и меристемните клетки.
При нормално осветление от тях директно се развиват останалите видове пластиди. Ако обаче диференцирането на пропластидите протича в отсъствие на светлина, те формират едно или няколко проламеларни телца (последните са с полукристална структура, изградена от тубуларни мембрани). Пропластиди съдържащи проламеларни телца се наричат етиопласти. Например, етиопласти се формират в клетките на листата на растения отглеждани на тъмно и в клетките на зародиша на семето. Когато растенията бъдат изложени на светло, етиопластите се трансформират в хлоропласти, като мембраните на проламеларните телца се развиват в тилакоиди. В клетките на зародиша на семето пропластидите се развиват първо в етиопласти и след поникването на младите растения и излагане на светлина, етиопластите се превръщат в хлоропласти.
Пластидите увеличават броя си чрез просто делене, подобно на бактериите. В меристемните клетки, пропластидите се делят с темпото на клетъчното делене. В останалите клетки, повечето пластиди водят началото си от делене на вече диференцираните пластиди.
Пластидите лесно могат взаимно да се превръщат едни в други, най-често в определена последователност (виж пунктираните стрелки на схемата)
Хлоропласти: органелите където протича фотосинтезата
 Хлоропластите са мястото където протича фотосинтезата - процес при който от СО2 и вода с участието на слънчевата светлина се синтезират въглехидрати. При висшите растения имат дисковидна форма с размери 4-6 μm в диаметър. Съдържат хлорофили (зелени пигменти) и каротиноиди (жълто-червени пигменти). Обикновено са ориентирани с широката си страна към стените на клетката, но могат да се преориентират под въздействие на светлината.
Хлоропластите са мястото където протича фотосинтезата - процес при който от СО2 и вода с участието на слънчевата светлина се синтезират въглехидрати. При висшите растения имат дисковидна форма с размери 4-6 μm в диаметър. Съдържат хлорофили (зелени пигменти) и каротиноиди (жълто-червени пигменти). Обикновено са ориентирани с широката си страна към стените на клетката, но могат да се преориентират под въздействие на светлината.
Растенията и другите еукариотни организми, които извършват фотосинтеза, обикновено съдържат в клетките си от един до няколко стотин хлоропласта. Една клетка от мезофила на листа най-често съдържа от 30 до 50 хлоропласта, а в 1mm² от листа се съдържат около 500 000 хлоропласта.
Хлоропластите дават очевидно предимство на организмите, които ги притежават - те могат да произвеждат своя собствена храна. Освен това те са абсолютния източник на всичката храна и горива които ние ползваме.
Освен в синтеза на въглехидрати, хлоропластите участват и в синтеза на аминокиселини и мастни киселини.
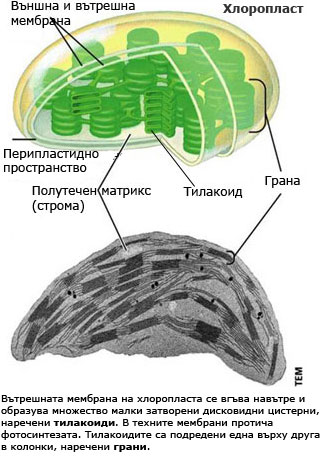
Структура на хлоропластите
Имат двойна мембрана и сложна вътрешна мембранна структура потопена в матрикс (строма). Вътрешната мембрана на хлоропласта се вгъва навътре и формира множество, малки, плоски, затворени дисковидни цистерни, наричани тилакоиди. Те са единна, взаимосвързана система и в мембраните им се намират фотосинтетичните пигменти, които директно участват във фотосинтезата. Тилакоидите са подредени в пакети (като колонка от монети) наречени грани. Всеки хлоропласт може да съдържа по сто и повече грани, а всяка грана е изградена от различен брой тилакоиди (2-100). Граните от своя страна са свързани помежду си посредством ламели на стромата. В матрикса се съдържат ензими, химични субстанции необходими за протичането на фотосинтезата, хлоропластна РНК, ДНК и рибозоми. Част от протеините, необходими за протичането на фотосинтезата се синтезират изцяло в матрикса на хлоропластите с участието на рибозомите намиращи се там, контролирани от хлоропластната ДНК. Но повечето от гените, които дефинират компонентите на хлоропластите се намират в ядрото на клетката.
Хлоропластите се образуват от пропластидите в меристемните клетки и увеличават броя си чрез просто делене в диференцираните клетки.
Лейкопластите са безцветни пластиди
Те са безцветните пластиди на клетката. Имат двойна обвивка, но не притежават пигменти и сложна вътрешна структура. В тях се синтезират и натрупват запасни хранителни вещества и се срещат в клетките на различните растителни органи. Според това дали в лейкопластите се отлагат скорбяла, протеини или мазнини, те съответно биват: амилопласти, протеопласти и олеопласти. 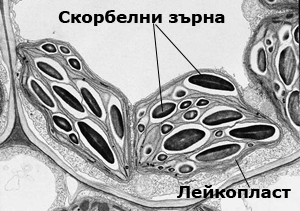 Амилопласти - в тях се извършва вторичната синтеза и натрупването на скорбяла, вследствие на което те се превръщат в скорбелни зърна. Скорбелните зърна могат да бъдат прости, когато скорбялата се отлага само около един център в лейкопласта и сложни, когато скорбялата се отлага около много центрове в лейкопласта. Скорбелните зърна са значително по-големи по размер. отколкото лейкопластите в които се образуват.
Амилопласти - в тях се извършва вторичната синтеза и натрупването на скорбяла, вследствие на което те се превръщат в скорбелни зърна. Скорбелните зърна могат да бъдат прости, когато скорбялата се отлага само около един център в лейкопласта и сложни, когато скорбялата се отлага около много центрове в лейкопласта. Скорбелните зърна са значително по-големи по размер. отколкото лейкопластите в които се образуват.
Протеопласти - в тях се натрупват протеини. Впоследствие обвивката и стромата на протеопласта се разрушават и в цитоплазмата се образуват малки белтъчни вакуоли.
Олеопласти - в тях под формата на малки глобули се отлагат мазнини. След като олеопластите бъдат изпълнен с мазнини, двойната обвивка и матриксът им се разрушават и в цитоплазмата на клетката се образуват маслени капки.
Хромопластите съдържат пигменти различни от хлорофил
Те са жълти, червени и оранжеви пластиди с 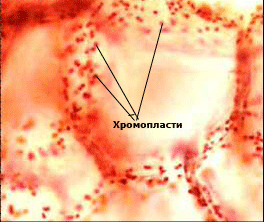 кръгла, ромбична, вретеновидна или игловидна форма. Имат двойна обвивка, полутечна строма и слаборазвита вътрешна мембранна система.
кръгла, ромбична, вретеновидна или игловидна форма. Имат двойна обвивка, полутечна строма и слаборазвита вътрешна мембранна система.
Пигментите им са от групата на каротиноидите:
- каротин (оранжево-червен)
- ксантофил (жълт)
- ликопин (червен)
Придават съответното оцветяване на много цветове, някои плодове, кореноплоди (морков).
Хромопластите се развиват от пропластиди, лейкопласти, млади хлоропласти или от дегенериране на напълно развити хлоропласти (хлорофилът и вътрешномембранната им структура се разграждат). Последният случай се наблюдава при узряването на много плодове (домати, пипер, тиква, шипка и др.)
Функция – привличат насекомите и животните и съдействат за опрашването на цветовете и разсейването на семената.
Цитоскелет: вътрешният скелет на клетката
Цитоплазмата на всички еукариотни клетки притежава мрежа от протеинови фибрили, наречена цитоскелет.
Цитоскелетът участва в много процеси като:
- делене, нарастване и диференциране на клетката,
- в движението на органелите в клетката,
- закотвя органелите в определени позиции,
- поддържа формата на клетката.
Той е динамична система, която постоянно се формира и дезинтегрира.
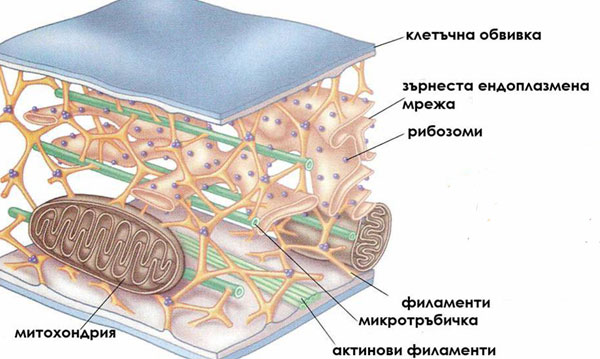
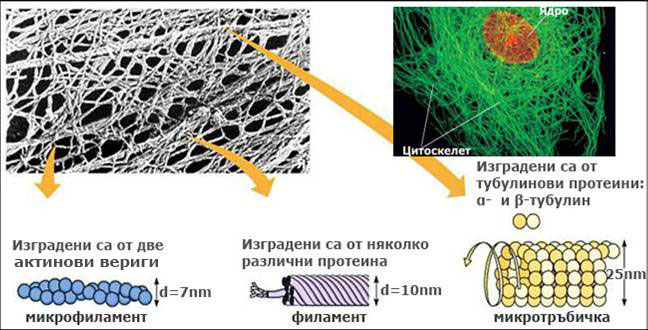
Три типа протеинови фибрили изграждат цитоскелета:
- Микрофиламенти. Наричат се още актинови филаменти и представляват дълги фибри с диаметър около 5-7 nm. Всеки микрофиламент е изграден от две дълги протеинови вериги, рехаво усукани заедно, подобно на две огърлици от перли. Всяка "перла" от огърлицата е субединица от глобуларния протеин актин. Актиновите филаменти се намират навсякъде в клетката, като се срещат поединично, асоциирани с микротръбичките или организирани в снопчета. Някои от тях, известни като стрес фибрили, са разположени близо до плазмената мембрана и се смята, че имат контрактилна функция. Други участват в прищъпването на животинските клетки при цитокинезата, контракцията на мускулните клетки, движението на органелите и цитоплазмата, изграждането на клетъчната стена и движението на транспортните мехурчета.
- Филаменти. Те са най-здравите елементи на цитоскелета и имат диаметър около 10 nm. Изградени са от различни протеинови фибри, които взаимно се застъпват и са силно усукани заедно. По този начин образуват въжеподобни структури с много голяма здравина. Обикновено веднъж формирани филаментите са стабилни и не се дезинтегрират. Малко се знае за ролята им в растителната клетка.
- Микротръбички. Представляват цилиндрични, кухи тръбички с диаметър около 25 nm. Всяка микротръбичка е изградена от субединици (димери) от протеините α- и β-тубулин, подредени в спирала. При изграждането на микротръбичките единият им край расте по-бързо от другия. Микротръбичките са динамични структури и преминават през регулярно дезинтегриране, формиране, ре-формиране и ре-аранжиране в нови конфигурации - състояние известно като динамична нестабилност. Изграждането на микротръбичките става в организационни центрове разположени на повърхността на ядрото и в кортикалната цитоплазма.
Микротръбичките участват в:
- организирането на вътреклетъчния транспорт в неделящите се клетки,
- ориентирането на целулозните микрофибрили при подреждането им в клетъчната стена;
- изграждането на делителното вретено и придвижването на хромозомите по време на деленето на клетката;
- в движението на органелите в клетката;
- изграждат камшичетата и ресничките и участват в движението на тези клетки.
Вакуоли: централният склад на растителните клетки
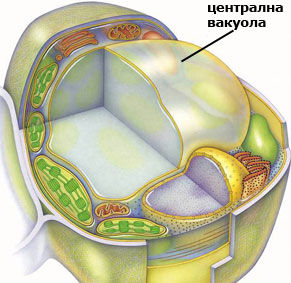 Вакуолите са изпълнени с клетъчен сок органели, отделени от останалата част на клетката с единична плазмена мембрана наречена тонопласт. Като правило в младите, растящи клетки има множество дребни вакуоли. С времето те увеличават размерите си и се сливат помежду си. Приключилите растежа си клетки, обикновено имат една голяма централна вакуола. Тя избутва цитоплазмата до плазмалемата и заема до 90% от обема на клетката.
Вакуолите са изпълнени с клетъчен сок органели, отделени от останалата част на клетката с единична плазмена мембрана наречена тонопласт. Като правило в младите, растящи клетки има множество дребни вакуоли. С времето те увеличават размерите си и се сливат помежду си. Приключилите растежа си клетки, обикновено имат една голяма централна вакуола. Тя избутва цитоплазмата до плазмалемата и заема до 90% от обема на клетката.
Вакуолите са мултифункционални органели и широко варират по размер, форма и съдържание на клетъчният им сок. Клетъчният сок представлява воден разтвор на различни неорганични соли, въглехидрати, аминокиселини, протеини, алкалоиди, органични киселини, гликозиди, дъбилни вещества, антоциани, пигменти и прочие. Вида на тези вещества варира и зависи освен от вида на растението, растителния орган и типа на клетките и от фазата на развитие и физиологичното състояние на растението.
Приема се, че вакуолите се образуват чрез уголемяване на сегменти на ендоплазмената мрежа или на мехурчета отделени от апарата на Голджи. Характерни са само за растителните клетки.
Функция на вакуолите
- Поддържат водния баланс и тургорното налягане на клетките.
- Служат като място за отлагане и на складиране на различни вещества. Химичните съединения акумулирани във вакуолите, по своята природа са или резервни за нуждите на клетката (най-вече първични метаболити) или токсични вторични метаболити изолирани там. Голяма част от вторичните метаболити натрупани във вакуолите са токсични не само за клетките, но и за патогените, паразитите и тревопасните животни и имат защитна функция за растенията.
Във вакуолите се натрупват и водоразтворими пигменти. Най-често това са антоциани, които придават син, лилав, тъмно червен, алено червен цвят на много плодове (грозде, сливи, череши), зеленчуци (зеле, репички), цветове (здравец, рози, божур, делфиниум, петуния). - Участват в разграждането и рециклирането на макромолекулите в клетката. Вакуолите, които съдържат големи количества хидролитични ензими участват в разграждането и рециклирането на макромолекулите в клетката. Дори цели органели, като остарели хлоропласти и митохондрии, могат да бъдат "погълнати" от вакуолите и разградени до съответните мономери. Тези т.н. литични вакуоли имат идентична функция с тази на лизозомите, характерни за животинските клетки.
Клетъчна стена
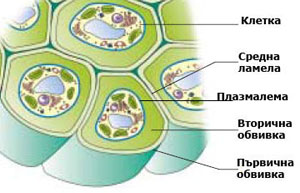 Клетъчната стена е плътна, многослойна структура с полизахаридна природа, която се отлага навън от плазмалемата (плазмената мембрана обграждаща клетката) на растителните клетки. Животинските клетки са ограничени само с плазмена мембрана.
Клетъчната стена е плътна, многослойна структура с полизахаридна природа, която се отлага навън от плазмалемата (плазмената мембрана обграждаща клетката) на растителните клетки. Животинските клетки са ограничени само с плазмена мембрана.
Функция на клетъчната стена
- Определя формата и размера на растителните клетки и допринася за дефиниране на окончателната форма на растителните органи.
- Противодейства на осмотичното налягане и не допуска разкъсването на плазмената мембрана в резултат на прекомерното поемане на вода.
- Определя механичната устойчивост на клетката и съответно и общата устойчивост на растенията – тя е твърдият скелет на клетката.
- Помага на растенията да противостоят на силите на гравитацията.
- Участва в абсорбирането, транспорта и секретирането на вещества.
- Предпазва клетката от инфекции посредством синтез на фитоалексини или синтез и депозиране на вещества (лигнин, кутин и др.), които представляват механична преграда пред гъби и бактерии.
- Свързва съседните клетки заедно.
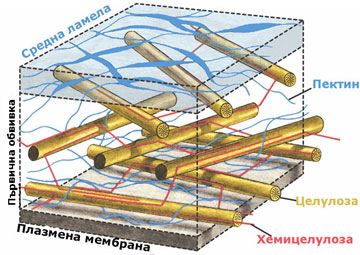
Химичен състав на клетъчната стена
Клетъчната стена е изградена от целулозни микрофибрили потопени в силно хидратиран матрикс от хемицелулози, пектинови вещества и малки количества гликопротеини и липиди.
Целулоза
Целулозата е основният компонент на клетъчната обвивка на растителните клетки и е най-разпространеният структурен полизахарид в растителния свят. 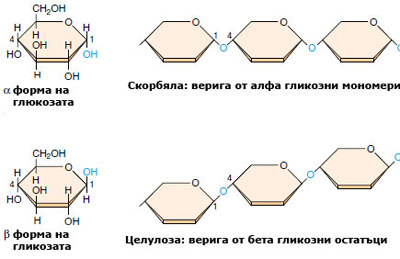 Тя е изградена от свързани помежду си, повтарящи се гликозни мономери. Характеризира се с изключителна механична и химична устойчивост. Неразтворима е във вода, слаби киселини и основи и обикновени органични разтворители. Империчната й формула е идентична с тази на скорбялата - (C6H10O5)n. Във веригата на скорбялата обаче всички гликозни мономери са α-гликоза, докато полимерните вериги на целулозата са изградени от β-гликозни единици. Тази разлика се оказва много съществена, понеже ензимите, които се срещат в повечето организми, лесно разграждат скорбялата, но не са в състояние да разрушават връзката между два β-гликозни мономера. И не защото тази връзка е по-здрава, а защото разрушаването й изисква ензим, който повечето организми не притежават. Изключителната здравина и устойчивост на целулозата я прави много добър биологичен структурен материал.
Тя е изградена от свързани помежду си, повтарящи се гликозни мономери. Характеризира се с изключителна механична и химична устойчивост. Неразтворима е във вода, слаби киселини и основи и обикновени органични разтворители. Империчната й формула е идентична с тази на скорбялата - (C6H10O5)n. Във веригата на скорбялата обаче всички гликозни мономери са α-гликоза, докато полимерните вериги на целулозата са изградени от β-гликозни единици. Тази разлика се оказва много съществена, понеже ензимите, които се срещат в повечето организми, лесно разграждат скорбялата, но не са в състояние да разрушават връзката между два β-гликозни мономера. И не защото тази връзка е по-здрава, а защото разрушаването й изисква ензим, който повечето организми не притежават. Изключителната здравина и устойчивост на целулозата я прави много добър биологичен структурен материал.
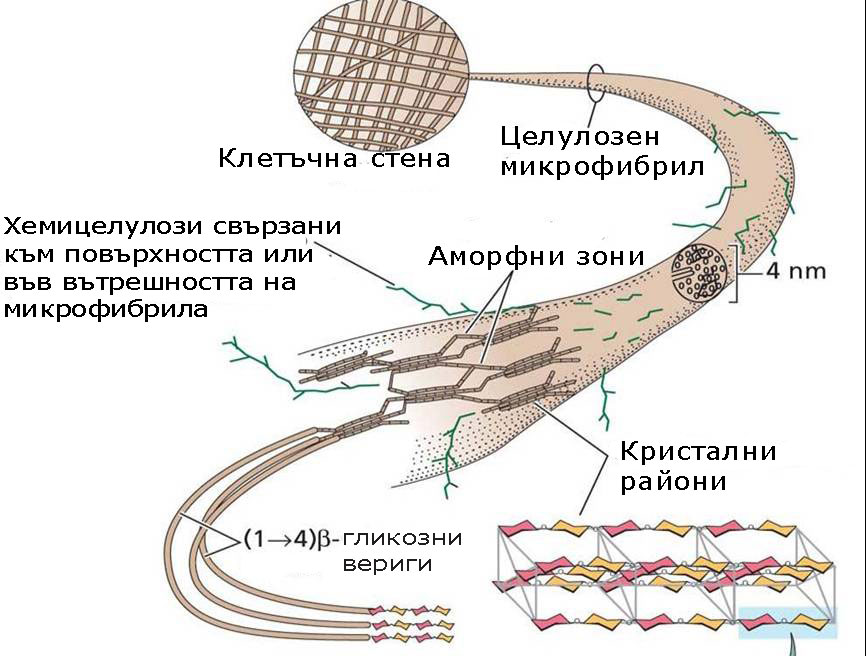 Молекулите на целулозата са дълги линейни вериги в които 300 до 3000 β-гликозни единици се свързват в една целулозна молекула. Тези дълги целулозни молекули имат свойството допълнително да се свързват помежду си чрез водородни връзки.
Молекулите на целулозата са дълги линейни вериги в които 300 до 3000 β-гликозни единици се свързват в една целулозна молекула. Тези дълги целулозни молекули имат свойството допълнително да се свързват помежду си чрез водородни връзки.
- 40-60 целулозни молекули разположени успоредно една на друга образуват елементарни фибрили или мицели, които ясно се виждат на електронен микроскоп.
- 15-20 елементарни фибрили организирани заедно образуват микрофибрили с диаметър от 4-10-25 nm.
- Целулозните микрофибрили от своя страна се усукват заедно, подобно на жичките в кабелите и формират макрофибрили с диаметър 0.5 µm, които лесно се наблюдават със светлинен микроскоп. Усукани по този начин целулозните макрофибрили имат здравина по-висока от тази на стоманени нишки със същата дебелина.
Целулозата има свойства на кристал и подобно на кристалите двойно пречупва светлинните лъчи. Това й свойство се наблюдава в мицелите - зоните където целулозните молекули са подредени успоредно една на друга и имат строга тримерна ориентация. Аморфните зони на микрофибрилите, където гликозни вериги са разположени около и между елементарните фибрили, не проявяват свойства на кристали.
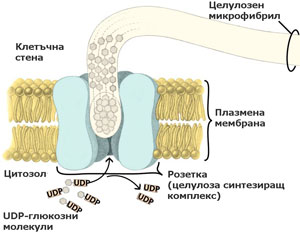 Целулозата се синтезира от целулоза синтезиращи комплекси локализирани в плазмалемата
Целулозата се синтезира от целулоза синтезиращи комплекси локализирани в плазмалемата
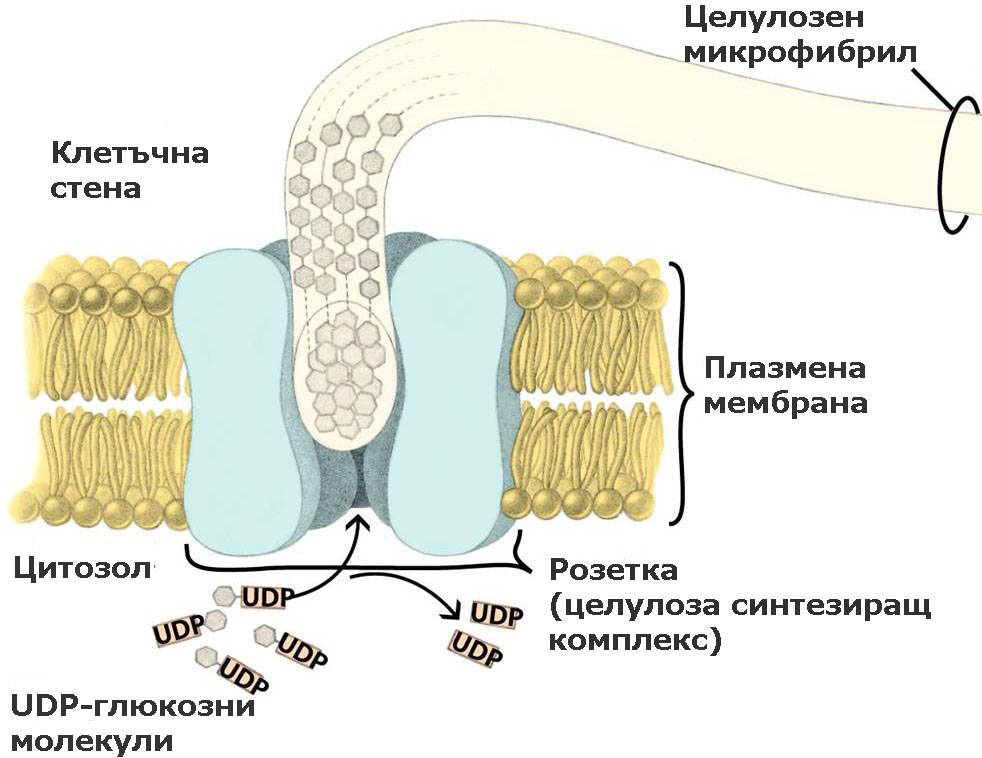
При семенните растения тези ензимни комплекси са организирани като пръстени или розетки от 6 единици, преминаващи през цялата широчина на плазмената мембрана. По време на целулозния синтез розетките се движат през плоскостта на плазмалемата и излъчват образуваните целулозни микрофибрили върху външната повърхност на плазмената мембрана. От там целулозните микрофибрили биват интегрирани в клетъчната стена. Приема се, че движението на розетките се регулира от микротръбички, разположени от вътрешната страна на плазмалемата.
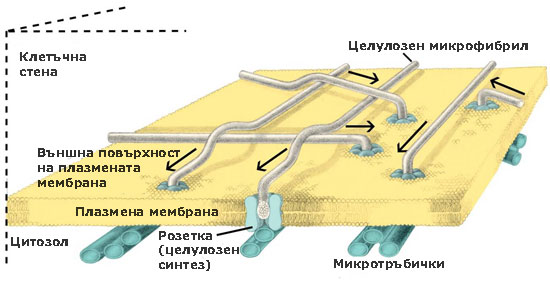 Единиците изграждащи розетките се доставят до и се вграждат в плазмената мембрана (плазмалемата) посредством транспортни мехурчета, образувани от апарата на Голджи.
Единиците изграждащи розетките се доставят до и се вграждат в плазмената мембрана (плазмалемата) посредством транспортни мехурчета, образувани от апарата на Голджи.
Матрикс на клетъчната стена
Матриксът на клетъчната стена е изграден от:
- хемицелулози,
- пектинови вещества и
- гликопротеини
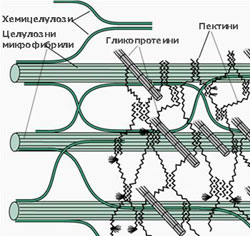 Хемицелулозите са полизахариди с дълги, неразклонени вериги, изградени основно от ксилоза и маноза и малки количества арабиноза и галактоза. Те не формират елементарни фибрили и химичният им състав варира силно, както при различните типове клетки, така и сред различните таксономични групи. Те ограничават еластичността на клетъчната стена, като се свързват посредством водородни връзки с целулозните микрофибрили и фиксират позициите им една спрямо друга.
Хемицелулозите са полизахариди с дълги, неразклонени вериги, изградени основно от ксилоза и маноза и малки количества арабиноза и галактоза. Те не формират елементарни фибрили и химичният им състав варира силно, както при различните типове клетки, така и сред различните таксономични групи. Те ограничават еластичността на клетъчната стена, като се свързват посредством водородни връзки с целулозните микрофибрили и фиксират позициите им една спрямо друга.
Пектините са калциеви и магнезиеви соли на галактуроновата киселина и арабинозата. Те имат силно разклонени молекули, без определена пространствена ориентация. Пектините изграждат изцяло средната ламела, която свързва съседните клетки и се съдържат в големи количества в първичната клетъчна обвивка (до 65%). Те са силно хигроскопични полизахариди и имат способността да поглъщат и задържат значителни количества вода.
Гликопротеини - те са или структурни протеини или ензими. Най-добре познати от структурните гликопротеини са екстензините, които заздравяват клетъчната стена и намаляват разширяването и еластичността й. В първичната клетъчна обвивка е открито и присъствието на голям брой ензими - пероксидази, фосфатази, целулази и пектинази.
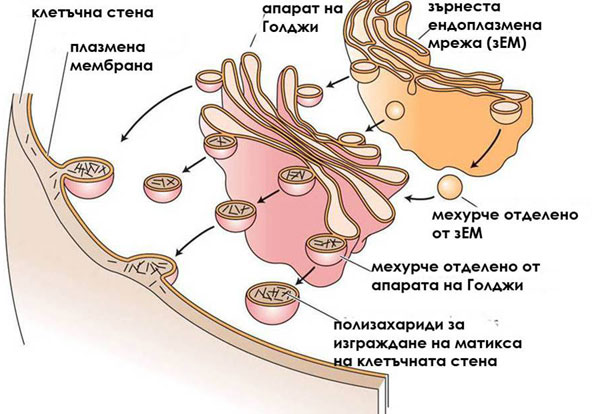
Компонентите на матрикса се синтезират от зърнестата ендоплазмена мрежа и апарата на Голджи
Хемицелулозите, пектините и гликопротеините се синтезират от зърнестата ендоплазмена мрежа и апарата на Голджи, след което се транспортират посредством транспортни мехурчета до плазмалемата. Там те се включват в изграждането на клетъчната обвивка. Типът на синтезираните компоненти на матрикса зависи от етапа на развитие на клетките. Например, пектините преобладават при растящите, удължаващи се клетки, докато хемицелулозите преобладават в клетките които са достигнали окончателните си размери.
Нарастването на клетъчната стена включва взаимодействие между плазмалемата, транспортните мехурчета и микротръбичките
Клетъчната обвивка нараства както на площ така и на дебелина. Нарастването й е комплексен процес, който протича под контрола на протопласта. При нарастването 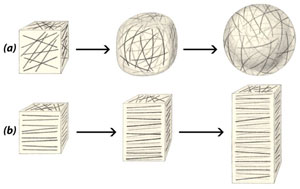 новообразуваните целулозни микрофибрили се отлагат, слой след слой, върху вече образуваните.
новообразуваните целулозни микрофибрили се отлагат, слой след слой, върху вече образуваните.
Начинът на ориентиране на целулозните микрофибрили в първичната клетъчна обвивка повлиява посоката на нарастване на клетките, съответно и формата, която придобиват. Ако целулозните микрофибрили във всички стени на клетката са ориентирани без определен порядък (a), тя придобива сферична форма. Ако целулозните микрофибрили се отлагат перпендикулярно на дългата ос на клетката (b), тя придобива удължена форма.
Много клетки образуват вторична клетъчна стена, в допълнение към първичната
Дебелината на клетъчните стени на растителните клетки варира силно в зависимост от (1) функцията, която те изпълняват и (2) в известна степен и от възрастта на отделните клетки.
Най-рано образуваните целулозни слоеве формират първичната клетъчна стена. Две съседни първични клетъчни стени се свързват със средна ламела (известна още като междуклетъчна субстанция). Много клетки образуват допълнителни слоеве клетъчна обвивка, които формират т.н. вторична клетъчна стена. Ако я има, вторичната клетъчна стена се синтезира и отлага върху повърхността на първичната.
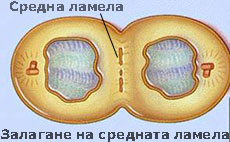 Средната ламела свързва съседните клетки
Средната ламела свързва съседните клетки
При митозата, цитоплазмата на делящата се майчина клетка се разделя с първична преграда, наречена клетъчна пластинка или средна ламела. Средната ламела е изградена основно от пектини и свързва новополучените две дъщерни клетки. Често е трудно да се разграничи средната ламела от първичната клетъчна стена, особено при клетки образуващи дебела вторична клетъчна стена. В тези случаи средната ламела, първичната клетъчна стена и евентуално и първия слой на вторичната клетъчна стена се наричат съставна средна ламела.
Първичната клетъчна стена се отлага докато клетките нарастват
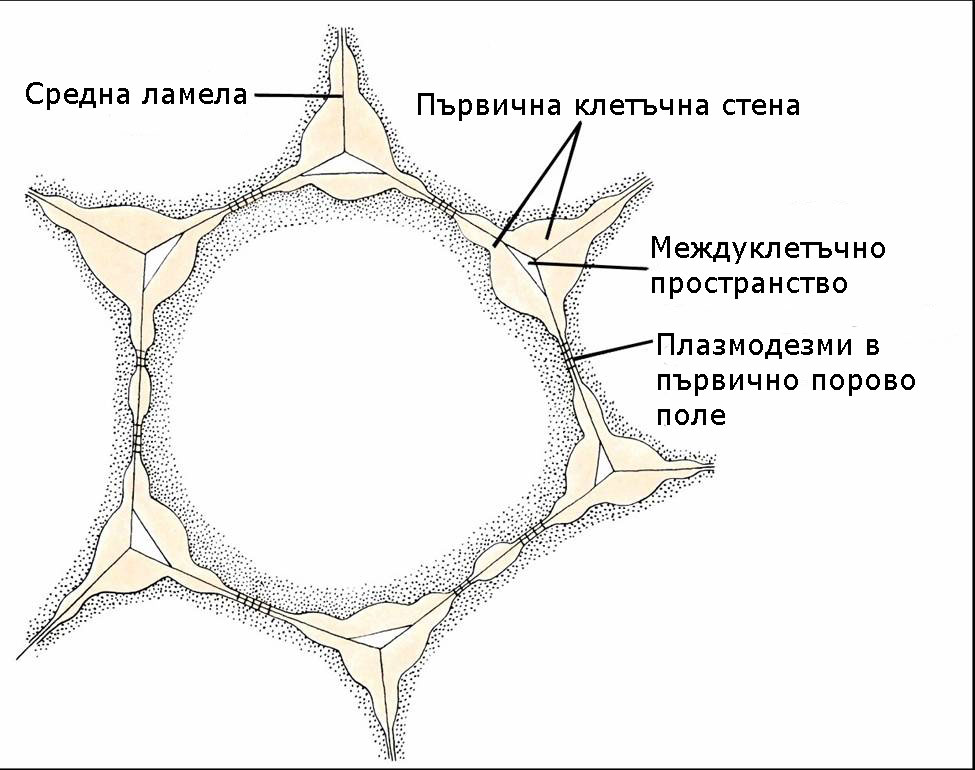 Първичната клетъчна обвивка е характерна за младите растящи растителни клетки. Тя е изградена главно от целулоза, хемицелулози, пектини, протеини (както гликопротеини така и ензими) и вода приблизително в следните съотношения:
Първичната клетъчна обвивка е характерна за младите растящи растителни клетки. Тя е изградена главно от целулоза, хемицелулози, пектини, протеини (както гликопротеини така и ензими) и вода приблизително в следните съотношения:
- целулоза (2-12%)
- хемицелулози (до 20%)
- пектинови вещества
- вода (до 60%)
Бидейки относително тънка и силно еластична, първичната клетъчна стена позволява безпрепятствено повърхностно нарастване на клетката.
Активно делящите се клетки, както и клетките участващи във фотосинтезата, дишането и секретирането на различни вещества, обикновено имат само първична клетъчна обвивка. Последните три вида клетки притежават способността да се дедиференцират и да придобиват способността да се делят отново, след което се диференцират в нов тип клетки. Поради това, принципно само живите клетки с първична клетъчна обвивка участват в заздравяването на раните и регенерирането на тъканите на растенията.
Първичните клетъчни стени също могат да съдържат лигнин, суберин и кутин.
Обикновено първичните клетъчни стени не са с еднаква дебелина по цялата си повърхност. Те имат по-тънки зони наричани порови полета. Плазмодезмите, които свързват протопласта на съседните клетки, обикновено са концентрирани в тези зони.
Вторичната клетъчна стена се отлага след приключване растежа на клетката и на първичната клетъчна стена
Някои клетки имат само първична клетъчна стена, но при други протопластът образува и вторична клетъчна стена. Тя се отлага навътре от първичната клетъчна стена и е локализирана между нея и плазмалемата. Вторичната клетъчна стена обикновено е характерна за специализирани клетки с механична или проводяща функция – коленхима, склеренхима, трахеи, трахеиди. Много от тези клетки умират след отлагането на вторичната клетъчна стена.
Целулозата е основен структурен материал във вторичната клетъчна стена, докато пектините са малко или дори могат да липсват. Това прави вторичната клетъчна стена твърда и неразтеглива. Матриксът й е изграден основно от хемицелулози. Структурните протеини и ензимите, които присъстват във първичната клетъчна стена, липсват във вторичната.
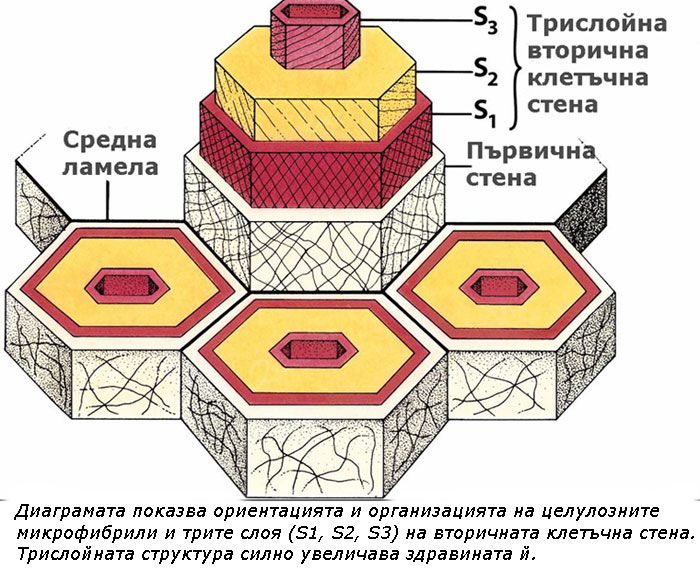 Често три слоя, обозначавани като S1, S2 S3 (за външния, средния и вътрешния слой съответно) могат да бъдат разграничени във вторичната клетъчна стена. Тези слоеве се различават един от друг по ориентацията на целулозните микрофибрили. Трислойната структура на вторичната клетъчна стена силно увеличава нейната здравина. Такива многослойни клетъчни стени се наблюдават при определени клетки на вторичната дървесина и обикновено съдържат и лигнин.
Често три слоя, обозначавани като S1, S2 S3 (за външния, средния и вътрешния слой съответно) могат да бъдат разграничени във вторичната клетъчна стена. Тези слоеве се различават един от друг по ориентацията на целулозните микрофибрили. Трислойната структура на вторичната клетъчна стена силно увеличава нейната здравина. Такива многослойни клетъчни стени се наблюдават при определени клетки на вторичната дървесина и обикновено съдържат и лигнин.
Където първичната клетъчна стена има порови полета, вторичната клетъчна стена има пори
При формирането си вторичната клетъчна стена не се отлага върху зоните, където са разположени поровите полета на първичната клетъчна стена. В резултат на това, характерни прекъсвания, известни като пори, се формират във вторичната стена. В някои случаи пори се формират и в зони където няма първични порови полета.
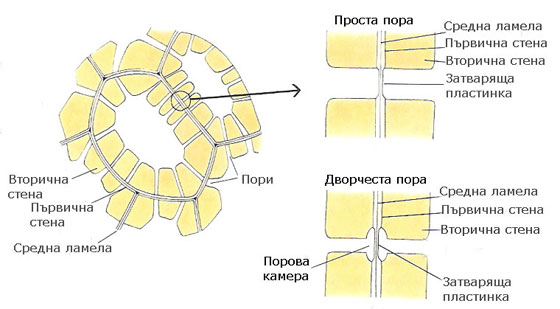
Порите на две съседните клетки обикновено възникват една срещу друга и формират двойки от пори. Средната ламела и двете първични клетъчни стени между две срещуположни пори се наричат затваряща пластинка.
Различаваме два основни типа пори - прости и дворчести
- При простите пори поровият канал е цилиндричен, елипсовиден или цепнатовиден; проекцията му е кръгче или елипса.
- При дворчестите пори се наблюдава надвесване на вторичната клетъчна обвивка над затварящата пластинка и поровият канал е като пресечен конус; проекцията е два вписани кръга.
Лигнифицираните вторични клетъчни стени са непропускливи за вода, но с формирането на порите, съседните клетки са разделени само чрез първичните клетъчни стени в зоната на затварящите им пластинки. Транспортът на веществата се извършва през затварящите пластинки на порите.
Отлагането на вторичната клетъчна стена се нарича още надебеляване на клетъчната стена
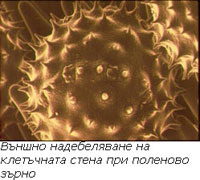
Надебеляването на клетъчната стена бива външно и вътрешно.
Външно надебеляване на клетъчната стена
Наблюдава се при клетки със свободна повърхност (спори, полен) и има формата на мрежи, шипчета, кукички и пр. То е типично за отделните видове и е важен систематичен белег.
Вътрешното надебеляване на клетъчната стена бива скулптурно и поресто
При скулптурното 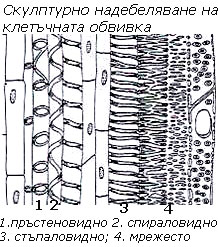 надебеляване надебелените участъци заемат по-малка част от повърхността на клетъчната обвивка и се отлагат под формата на някаква фигура (скулптура). Според формата на скулптурите този вид надебеляване бива:
надебеляване надебелените участъци заемат по-малка част от повърхността на клетъчната обвивка и се отлагат под формата на някаква фигура (скулптура). Според формата на скулптурите този вид надебеляване бива:
- пръстеновидно,
- спираловидно,
- стъпаловидно и
- мрежесто
При порестото надебеляване на клетъчната стена надебелените места заемат по-голяма част от обвивката и само малки участъци, наричани пори, остават ненадебелени. Транспортът на веществата се извършва през порите.
Плазмодезмите са цитоплазмени нишки, които свързват протопласта на съседните клетки
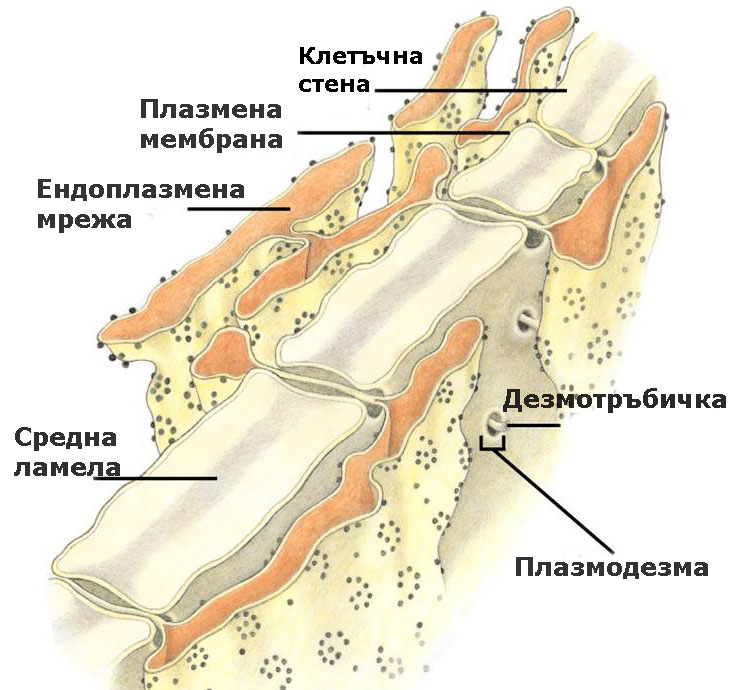 Цитоплазмата на съседните клетки е свързана посредством плазмодезми, които обикновено преминават през първичните порови полета или порите на клетъчните стени и участват в транспорта на веществата и в предаването на сигнали между клетките.
Цитоплазмата на съседните клетки е свързана посредством плазмодезми, които обикновено преминават през първичните порови полета или порите на клетъчните стени и участват в транспорта на веществата и в предаването на сигнали между клетките.
Под електронен микроскоп, те се наблюдават като тесни каналчета с диаметър от 30-60 nm.
Плазмодезмите са тесни каналчета, покрити с плазмена мембрана и изпълнени с цитозол и разположена в средата дезмотръбичка.
Дезмотръбичката представлява модифицирана, тясна и плътна тръбичка на ЕМ, без лумен. Целия транспорт на веществата става през обграждащия я цитозол.
Понеже клетките са свързани посредством плазмодезмите, протопластът на цялото растение, представлява единно цяло и се нарича симпласт. Съответно, движението на веществата през плазмодезмите, се нарича транспорт на веществата по симпласта.
От друга страна, клетъчните стени също образуват единно цяло, наричано апопласт. Движението на веществата по клетъчните стени, се нарича транспорт на веществата по апопласта.
Повечето плазмодезми се формират по време на делене на клетката, като нишки от 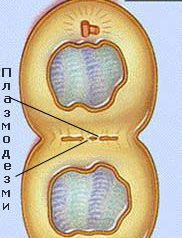 ендоплазмената мрежа, се оказват оградени от залагащата се средна ламела. Плазмодезми могат да се залагат и de novo във вече формираните клетъчни стени. Плазмодезмите формирани през цитокинезата се наричат първични плазмодезми, а тези формирани след цитокинезата - вторични плазмодезми.
ендоплазмената мрежа, се оказват оградени от залагащата се средна ламела. Плазмодезми могат да се залагат и de novo във вече формираните клетъчни стени. Плазмодезмите формирани през цитокинезата се наричат първични плазмодезми, а тези формирани след цитокинезата - вторични плазмодезми.
Броят на плазмодезмите варира и може да достигне до 20 000 в клетка.
Изменения в химичния състав на клетъчните стени
В процеса на развитие на клетката, клетъчната й стена се изменя съществено. Тези промени могат да се дължат на промени в основните компоненти, които изграждат клетъчната обвивка или на допълнително отлагане на нови химични съединения. Новите вещества могат да се отлагат в матрикса на клетъчната стена между целулозните микрофибрили (процеса се нарича инкрустиране) или да се отлагат върху целулозните микрофибрили (процеса се нарича адкрустиране).
Лигнифициране или вдървесиняване на клетъчната стена
 Изразява се в отлагането на лигнини в клетъчните стени. Лигнините са фенолни полимери, изключително широко разпространени в природата - втори след целулозата. Съотношението на мономерите, които ги изграждат (p-кумарил, кониферил и синапил алкохоли), варира значително, както при различните голосеменни и покритосеменни растения, така и при отделните растителни видове, органи и тъкани. Лигнините имат следната роля:
Изразява се в отлагането на лигнини в клетъчните стени. Лигнините са фенолни полимери, изключително широко разпространени в природата - втори след целулозата. Съотношението на мономерите, които ги изграждат (p-кумарил, кониферил и синапил алкохоли), варира значително, както при различните голосеменни и покритосеменни растения, така и при отделните растителни видове, органи и тъкани. Лигнините имат следната роля:
- Придават твърдост и здравина на клетъчните стени. Смята се, че са изиграли основна роля в еволюцията на сухоземните растения. Отлагането на лигнини е дало необходимата механична устойчивост на растенията да противодействат на силите на гравитацията и да развият изправени и силно разклонени стъбла, способни да поддържат големи фотосинтетични повърхности.
- Правят клетъчните стени водонепропускливи и подпомагат възходящия транспорт на водата в проводящите елементи на ксилема.
- Защитават растенията. При нападения от гъби и бактерии или при наранявания се наблюдава отлагане на лигнин в съответните зони. Така наречения раневи лигнин предпазва растението, като повишава здравината на клетъчните стени и те противодействат, както на физическото проникване на гъбите в растителните тъкани, така и на дифузията на ензимите и токсините отделяни от тях. Някои учени смятат, че при еволюцията на сухоземните растения, лигнините първоначално са действали като антибактериални и антигъбни агенти и едва по-късно са придобили съществена роля в транспорта на веществата и повишаване на механичната устойчивост на растенията.
Лигнифицирането на клетъчните стени обикновено води до загиване на протопласта и се наблюдава основно при проводящите и механичните тъкани на растенията.
 Вкорковяване или суберинизиране на клетъчната стена
Вкорковяване или суберинизиране на клетъчната стена
Представлява адкрустиране на суберин в клетъчната обвивка. Суберинът е липидно съединение отличаващо се с голяма химична устойчивост. Вкорковените клетъчни стени не пропускат вода и газове и протопласта загива. Суберинът е главен компонент на стените на корковите клетки, изграждащи най-външния слой на кората на растенията. На електронен микроскоп в суберинизираните клетъчни стени се наблюдават редуващи се светли и тъмни слоеве. Смята се, че светлите слоеве са изградени от восъци, а тъмните - от суберин.
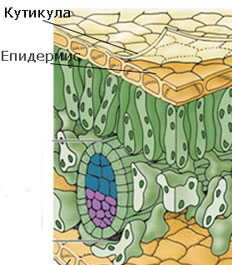 Кутинизация на клетъчната стена
Кутинизация на клетъчната стена
Кутинът е липидно вещество, което е важен структурен компонент на клетъчните стени на много растения. Основната му функция е да формира матрикс, в който се разполагат восъци (липидни съединения с високо молекулно техло и дълги вериги). Восъците в комбинация с кутин формират защитен слой върху външните стени на епидермалните клетки, известен като кутикула. Освен восъци вградени в кутина (кутикулярни восъци), кутикулата често е покрита и от повърхностен слой от восъци (епикутикулярни восъци). Кутикулата (1) предотвратява загубата на вода и други молекули от повърхността на растителните органи и (2) предпазва растенията и от бактериални и гъбни заболявания, като не позволява задържането на вода върху растителните органи.
Мацерация
Представлява разединяване на клетките, вследствие на разрушаване на свързващата ги средна ламела. Бива естествена и изкуствена. Естествената мацерация протича под въздействието на ензима пектиназа, който разгражда пектиновите вещества на средната ламела. Причинява окапването на листата, венчелистчетата, узрелите плодове, както и отделянето на техническите влакна на лена и конопа. Изкуствена мацерация може да бъде предизвикана чрез въздействие с висока температура, синтетични ензими, разредени киселини. При варенето на кореноплоди, плодове, картофи и други продукти от растителен произход винаги настъпва мацерация, в резултат на което те омекват.
 Ослизяване на клетъчната стена
Ослизяване на клетъчната стена
Слизестите вещества представляват хетерополизахариди със способност силно да набъбват във вода. Те се синтезират от растенията с цел поглъщане и задържане на вода. Получават се чрез трансформиране на целулоза, скорбяла и пектинови вещества и се локализират в определени клетки в растителните тъкани. Понякога слизестите вещества се отлагат направо върху клетъчната обвивка като вторично надебеляване – явление което се наблюдава при семена на лен и дюля. Слизестите вещества подобряват контакта на семената с почвените частици, а също поглъщат и задържат на вода, с което улесняват покълването на семената.
Гумозис на клетъчната стена
 Гумите са хетерополизахариди и обикновено се образуват в резултат на нараняване на кората на дърветата. Процесът е известен като “гумозиc” и при него клетъчните стени, цели клетки, а понякога и цели групи от клетки в зоната на нараненото място се втечняват и формират вискозен, лепкав, колоиден разтвор. Този колоиден разтвор формира защитен покривен слой върху наранената тъкан и не позволява проникване на гъби и бактерии през нараненото място. Впоследствие гумозните вещества изсъхват и се втвърдяват по повърхността на кората като различни по форма и цвят полупрозрачни зърна или късчета. Гумозис се наблюдава при слива, джанка, череша, бадем и др. При някои растения, синтезът на гумите може да стане и без патологична причина, като част от механизъм за задържане на вода. Наблюдава се при растения развиващи се в сухи местообитания и освен в кората може да протича и в други части на растението - корени, семена, плодове.
Гумите са хетерополизахариди и обикновено се образуват в резултат на нараняване на кората на дърветата. Процесът е известен като “гумозиc” и при него клетъчните стени, цели клетки, а понякога и цели групи от клетки в зоната на нараненото място се втечняват и формират вискозен, лепкав, колоиден разтвор. Този колоиден разтвор формира защитен покривен слой върху наранената тъкан и не позволява проникване на гъби и бактерии през нараненото място. Впоследствие гумозните вещества изсъхват и се втвърдяват по повърхността на кората като различни по форма и цвят полупрозрачни зърна или късчета. Гумозис се наблюдава при слива, джанка, череша, бадем и др. При някои растения, синтезът на гумите може да стане и без патологична причина, като част от механизъм за задържане на вода. Наблюдава се при растения развиващи се в сухи местообитания и освен в кората може да протича и в други части на растението - корени, семена, плодове.
 Минерализиране на клетъчната стена
Минерализиране на клетъчната стена
Представлява инкрустиране на различни минерални вещества в клетъчните стени. Най-често това са силициев двуокис и калциев карбонат. Минерализирането на клетъчните стени прави растителните органи твърди, грапави и намалява фуражната им стойност. Има защитна функция за растенията, като ги предпазва от неприятели и тревопасни животни. Наблюдава се при хвощовете, житните растения, видовете от сем. Boraginaceae (Грапаволистни) и сем. Brassicaceae (Зелеви) и др.